PUFA stabilizes a conductive state of the selectivity filter in IKs channels
Curation statements for this article:-
Curated by eLife

eLife assessment
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating and PUFA modulation of this mechanism. This mechanism is supported by convincing single channel recordings, macroscopic current recordings and mutational analyses. These findings are of importance to the ion channel field and possibly future therapeutic applications.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
- Reading List (BiophysicsColab)
Abstract
In cardiomyocytes, the KCNQ1/KCNE1 channel complex mediates the slow delayed-rectifier current (IKs), pivotal during the repolarization phase of the ventricular action potential. Mutations in IKs cause long QT syndrome (LQTS), a syndrome with a prolonged QT interval on the ECG, which increases the risk of ventricular arrhythmia and sudden cardiac death. One potential therapeutical intervention for LQTS is based on targeting IKs channels to restore channel function and/or the physiological QT interval. Polyunsaturated fatty acids (PUFAs) are potent activators of KCNQ1 channels and activate IKs channels by binding to two different sites, one in the voltage sensor domain – which shifts the voltage dependence to more negative voltages – and the other in the pore domain – which increases the maximal conductance of the channels (Gmax). However, the mechanism by which PUFAs increase the Gmax of the IKs channels is still poorly understood. In addition, it is unclear why IKs channels have a very small single-channel conductance and a low open probability or whether PUFAs affect any of these properties of IKs channels. Our results suggest that the selectivity filter in KCNQ1 is normally unstable, contributing to the low open probability, and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state.
Article activity feed
-

-
-
-

eLife assessment
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating and PUFA modulation of this mechanism. This mechanism is supported by convincing single channel recordings, macroscopic current recordings and mutational analyses. These findings are of importance to the ion channel field and possibly future therapeutic applications.
-
Reviewer #1 (Public review):
This study comes to an interesting conclusion: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state is well supported by single channel analysis, which shows that normally infrequent opening bursts occur more often in the presence of the PUFA. The linkage to PUFA action through the selectivity filter is supported by disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. A definitive functional experiment is conducted by single channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. The computational exploration of two selectivity …
Reviewer #1 (Public review):
This study comes to an interesting conclusion: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state is well supported by single channel analysis, which shows that normally infrequent opening bursts occur more often in the presence of the PUFA. The linkage to PUFA action through the selectivity filter is supported by disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. A definitive functional experiment is conducted by single channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. The computational exploration of two selectivity filter structures proposed to interact distinctly with Lin-Glycine is informative. Both mutation results and simulations converge on the proposed selectivity filter mechanism, although other possibilities for Lin-Glycine binding and action might be possible. Overall, the major claim of the abstract is well-supported: "... that the selectivity filter in KCNQ1 is normally unstable ... and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state."
-
Reviewer #2 (Public review):
Golluscio et al. address one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by polyunsaturated fatty acids (PUFAs). PUFAs are known to upregulate KCNQ1 and KCNQ1/KCNE1 channels through two mechanisms: one shifts the voltage dependence in a negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium through a charge effect, the second mechanism is less understood. Using single-channel recordings and mutagenesis at putative PUFA binding sites, they successfully demonstrate that the selectivity filter is stabilized in a conducting state by PUFA binding, and that this is the mechanism by which PUFAs increase Gmax. Their single-channel recordings are straightforward and clearly show that the selectivity filter …
Reviewer #2 (Public review):
Golluscio et al. address one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by polyunsaturated fatty acids (PUFAs). PUFAs are known to upregulate KCNQ1 and KCNQ1/KCNE1 channels through two mechanisms: one shifts the voltage dependence in a negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium through a charge effect, the second mechanism is less understood. Using single-channel recordings and mutagenesis at putative PUFA binding sites, they successfully demonstrate that the selectivity filter is stabilized in a conducting state by PUFA binding, and that this is the mechanism by which PUFAs increase Gmax. Their single-channel recordings are straightforward and clearly show that the selectivity filter tends to become conductive upon PUFA binding. Since PUFAs are potential therapeutic reagents for cardiac arrhythmias such as long QT syndrome, their findings are beneficial for future research and applications of these compounds.
-
Reviewer #3 (Public review):
Summary:
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating such that the open state of the pore is unstable and undergoes intermittent closed and open conformations. PUFA enhances the maximum open probability of IKs by binding to a crevice adjacent to the pore and stabilize the open conformation. This mechanism is supported by convincing single channel recordings that show empty and open channel traces and the ratio of such traces is affected by PUFA. In addition, mutations of the pore residues alter PUFA effects, convincingly supporting that PUFA alters the interactions among these pore residues.
Strengths:
The data are of high quality and the description is clear.
-
Author response:
The following is the authors’ response to the previous reviews.
Reviewer #1 (Recommendations For The Authors):
The additional data included in this revision nicely strengthens the major claim.
I apologize that my comment about K+ concentration in the prior review was unclear. The cryoEM structure of KCNQ1 with S4 in the resting state was obtained with lowered K+ relative to the active state. Throughout the results and discussion it seems implied that the change in voltage sensor state is somehow causative of the change in selectivity filter state while the paper that identified the structures attributes the change in selectivity filter state not to voltage sensors, but to the change in [K+] between the 2 structures. Unless there is a flaw in my understanding of the conditions in which the selectivity filter structures …
Author response:
The following is the authors’ response to the previous reviews.
Reviewer #1 (Recommendations For The Authors):
The additional data included in this revision nicely strengthens the major claim.
I apologize that my comment about K+ concentration in the prior review was unclear. The cryoEM structure of KCNQ1 with S4 in the resting state was obtained with lowered K+ relative to the active state. Throughout the results and discussion it seems implied that the change in voltage sensor state is somehow causative of the change in selectivity filter state while the paper that identified the structures attributes the change in selectivity filter state not to voltage sensors, but to the change in [K+] between the 2 structures. Unless there is a flaw in my understanding of the conditions in which the selectivity filter structures used in modeling were generated, it seems misleading to ignore the change in [K+] when referring to the activated vs resting or up vs down structures. My understanding is that the closed conformation adopted in the resting/low [K+] is similar to that observed in low [K+] previously and is more commonly associated with [K+]-dependent inactivation, not resulting from voltage sensor deactivation as implied here. The original article presenting the low [K+] structure also suggests this. When discussing conformational changes in the selectivity filter, I strongly suggest referring to these structures as activated/high [K+] vs resting/low [K+] or something similar, as the [K+] concentration is a salient variable.
There seems to be some major confusion here and we will try to explain how we think. Note that in the Mandela and MacKinnon paper, there is no significant difference in the amino acid positions in the selectivity filter between low and high K+ when S4 is in the activated position (See Mandala and Mackinnon, PNAS Suppl. Fig S5 C and D). There are only fewer K+ in the selectivity filter in low K+. So, the structure with the distorted selectivity filter is not due to low K+ by itself. Note that there is no real difference between macroscopic currents recorded in low and high K+ solutions (except what is expected from changes in driving force) for KCNQ1/KCNE1 channels (Larsen et al., Bioph J 2011), suggesting that low K+ do not promote the non-conductive state (Figure 1). We now include a section in the Discussion about high/low K+ in the structures and the absence of effects of K+ on the function of KCNQ1/KCNE1 channels.
Author response image 1.
Macroscopic KCNQ1/KCNE1 currents recorded in different K+ conditions. Note that there is no difference between current recorded in low K+ (2 mM) conditions and high (96 mM) K+ conditions (n=3 oocytes). Currents were normalized in respect to high K+.
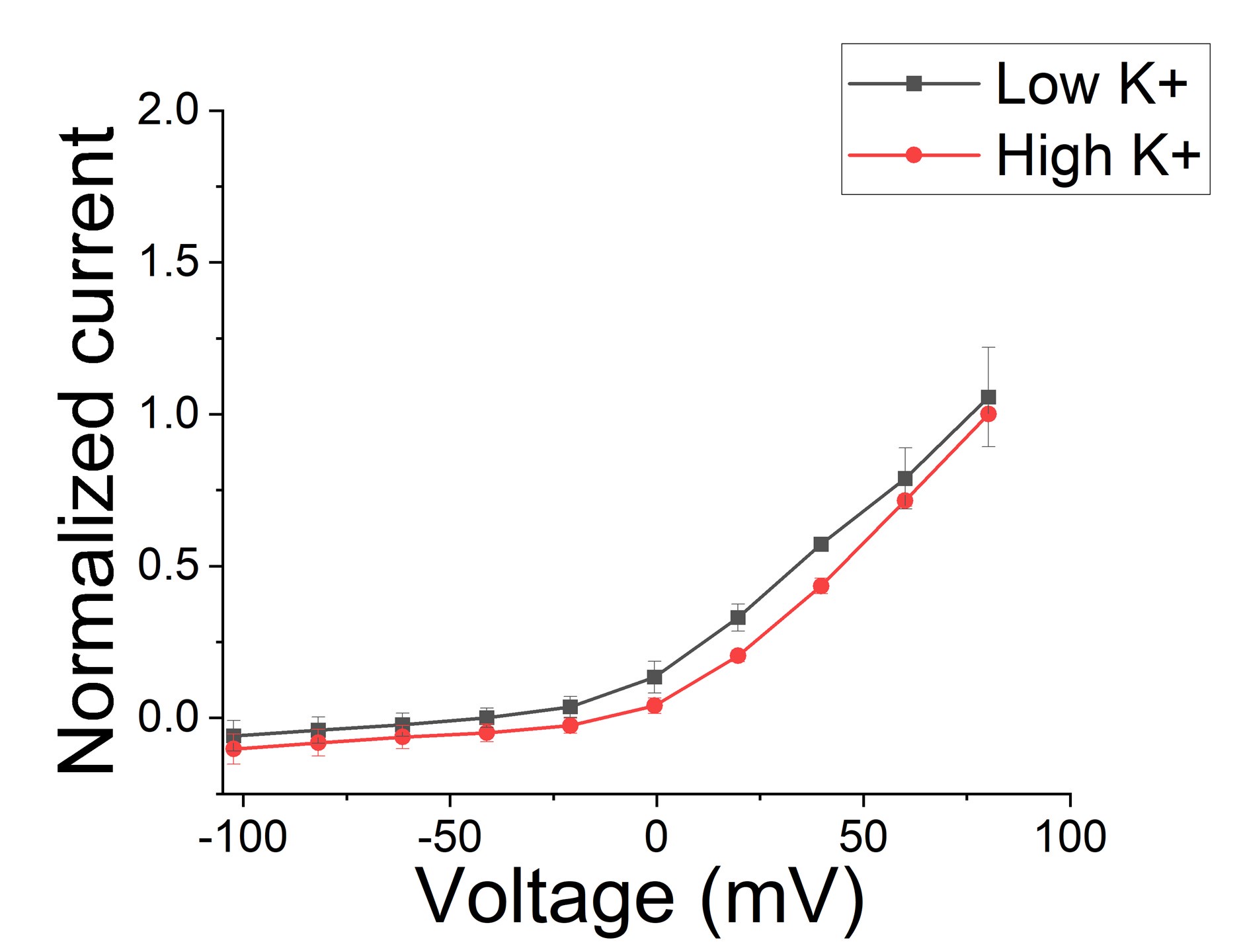
Note also that, in the previous version of the manuscript, we did not propose that the position of S4 is what determines the state of the selectivity filter. We only reported that the CryoEM structure with S4 resting shows a distorted selectivity filter. It seems like our text confused the reviewer to think that we proposed that S4 determines the state of the selectivity filter, when we did not propose this earlier. We previously did not want to speculate too much about this, but we have now included a section in the Discussion to make our view clear in light of the confusion of the reviewers.
It is clear from our data that the majority of sweeps are empty (which we assume is with S4 up), suggesting that the selectivity filter can be (and is in the majority of sweeps) in the non-conducting state even with S4 up. We think that the selectivity filter switches between a non-conductive and a conductive conformation both with S4 down and with S4 up. The cryoEM structure in low K+ and S4 down just happened to catch the non-conductive state of the selectivity filter. We have now added a section in the Discussion to clarify all this and explain how we think it works.
However, S4 in the active conformation seems to stabilize the conductive conformation of the selectivity filter, because during long pulses the channel seems to stay open once opened (See Suppl Fig S2). So, one possibility is that the selectivity filter goes more readily into the non-conductive state when S4 is down (and maybe, or not, low K+ plays a role) and then when S4 moves up the selectivity filter sometimes recovers into the conductive state and stays there. We now have included a section in the Discussion to present our view. Since this whole discussion was initiated and pushed by the reviewer, we hope that the reviewers will not demand more data to support these ideas. We think that this addition makes sense since other readers might have the same questions and ideas as the reviewer, and we would like to prevent any confusion about this topic.
Figure 1
It remains unclear in the manuscript itself what "control" refers to. Are control patched the same patches that later receive LG?
Yes, the control means the same patch before LG. We now indicate that in legends and text throughout.
Supplementary Figure S1
Unclear if any changes occur after addition of LG in left panel and if the LG data on right is paired in any way to data on left.
Yes, in all cases the left and right panel in all figures are from the same patch. We now indicate that in legends and text throughout.
The letter p is used both to represent open probability open probability from the all-point amplitude histogram and as a p-value statistical probability indicator sometime lower case, sometimes upper case. This was confusing.
We have now exclusively use lower case p for statistical probability and Po for open probability.
"This indicates that mutations of residues in the more intracellular region of the selectivity filter do not affect the Gmax increases and that the interactions that stabilize the channel involve only residues located near the external region part of the selectivity filter. "
Seems too strongly worded, it remains possible that mutations of other residues in the more intracellular region of the selectivity filter could affect the Gmax increases.
We have changed the text to: "Mutations of residues in the more intracellular region of the selectivity filter do not affect the Gmax increases, as if the interactions that stabilize the channel involve residues located near the external region part of the selectivity filter. "
Supplementary Figure S7
Please report Boltzmann fit parameters. What are "normalized" uA?
We removed the uA, which was mistakenly inserted. The lines in the graphs are just lines connecting the dots and not Boltzmann fits, since we don’t have saturating curves in all panels to make unique fits.
"We have previously shown that the effects of PUFAs on IKs channels involve the binding of PUFAs to two independent sites." Was binding to the sites actually shown? Suggest changing to: "We have previously proposed models in which the effects of PUFAs..."
We have now changed this as the Reviewer suggested: " We have previously proposed models in which the effects of PUFAs on IKs channels involve the binding of PUFAs to two independent sites."
Statistics used not always clear. Methods refer to multiple statistical tests but it is not clear which is used when.
We use two different tests and it is now explained in figure legends when either was used.
n values confusing. Sometimes # of sweeps used as n. Sometimes # patches used as n. In one instance "The average current during the single channel sweeps was increased by 2.3 {plus minus} 0.33 times (n = 4 patches, p =0.0006)" ...this sems a low p value for this n=4 sample?
We have now more clearly indicated what n stands for in each case. There was an extra 0 in the p value, so now it is p = 0.006. Thanks for catching that error.
Reviewer #2 (Recommendations For The Authors):
I still have some comments for the revised manuscript.
(1) (From the previous minor point #6) Since D317E and T309S did not show statistical significance in Figure 5A, the sentences such as "This data shows that Y315 and D317 are necessary for the ability of Lin-Glycine to increase Gmax" or "the effect of Lin-Glycine on Gmax of the KCNQ1/KCNE1 mutant was noticeably reduced compared to the WT channel showing the this residue contributes to the Gmax effect (Figure 5A)." may need to be toned down. Alternatively, I suggest the authors refer to Supplementary Figure S7 to confirm that Y315 and D317 are critical for increasing Gmax.
We have redone the analysis and statistical evaluation in Fig 5. We no use the more appropriate value of the fitted Gmax (which use the whole dose response curve instead of only the 20 mM value) in the statistical evaluation and now Y315F and D317E are statistically different from wt.
(2) Supplementary Fig. S1. All control diary plots include the green arrows to indicate the timing of lin-glycine (LG) application. It is a bit confusing why they are included. Is it to show that LG application did not have an immediate effect? Are the LG-free plots not available?
Not sure what the Reviewer is asking about? In the previous review round the Reviewers asked specifically for this. The arrow shows when LG was applied and the plot on the right shows the effect of LG from the same patch.
(3) The legend to Supplementary Figure S4, "The side chain of residues ... are highlighted as sticks and colored based on the atomic displacement values, from white to blue to red on a scale of 0 to 9 Å." They look mostly blue (or light blue). Which one is colored white? It might be better to use a different color code. It would also be nice to link the color code to the colors of Supplementary Figure S5, which currently uses a single color.
We have removed “from white to blue to red on a scale of 0 to 9 Å” and instead now include a color scale directly in Fig S4 to show how much each atom moved based on the color.
We feel it is not necessary to include color in Fig S5 since the scale of how much each atom moves is shown on the y axis.
(4) Add unit (pA) to the y-axis of Supplementary Figure S2.
pA has been added.
Reviewer #3 (Recommendations For The Authors):
Some issues on how data support conclusions are identified. Further justifications are suggested.
186: “The decrease in first latency is most likely due to an effect of Lin-Glycine on Site I in the VSD and related to the shift in voltage dependence caused by Lin-Glycine." The results in Fig S1B do not seem to support this statement since the mutation Y315F in the pore helix seemed to have eliminated the effect of Lin-Glycine in reducing first latency. The authors may want to show that a mutation that eliminating Site I would eliminate the effect of Lin-Glycine on first latency. On the other hand, it will be also interesting to examine if another pore mutation, such as P320L (Fig 5) also reduce the effect of Lin-Glycine on first latency.
These experiments are very hard and laborious, and we feel these are outside the scope of this paper which focuses on Site II and the mechanism of increasing Gmax. Further studies of the voltage shift and latency will have to be for a future study.
The mutation D317E did not affect the effect of Lin-Glycine on Gmax significantly (Fig 5A, and Fig S7F comparing with Fig S7A), but the authors conclude that D317 is important for Lin-Glycine association. This conclusion needs a better justification.
We have redone the analysis and statistical evaluation in Fig 5. We no use the more appropriate value of the fitted Gmax (which use the whole dose response curve instead of only the 20 mM value) in the statistical evaluation and now D317E is statistically different from wt
-
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single-channel analysis, though some method details are missing. The linkage to PUFA action through the selectivity filter is supported by the disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. Claims about differences in Lin-Glycine binding to these two structural conformations seem to lack clear support, thus the claim seems speculative that PUFAs increase Gmax by …
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single-channel analysis, though some method details are missing. The linkage to PUFA action through the selectivity filter is supported by the disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. Claims about differences in Lin-Glycine binding to these two structural conformations seem to lack clear support, thus the claim seems speculative that PUFAs increase Gmax by binding to a crevice in the pore domain. A potentially definitive functional experiment is conducted by single-channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. However, this appears to be an n=1 experiment. Overall, the major claim of the abstract is supported: "... that the selectivity filter in KCNQ1 is normally unstable ... and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state." However, the claim in the abstract that selectivity filter instability "explains the low open probability" seems too general.
We thank the reviewer for the comments, and we would like to address the main concern regarding the single channels. We now state the number of experiments used for the single channel analysis. We agree that the claim in the abstract seems too general and we now made it more specific to our findings.
Reviewer #2 (Public Review):
Golluscio et al. address one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by polyunsaturated fatty acids (PUFA). PUFA is known to upregulate KCNQ1 and KCNQ1/KCNE1 channels by two mechanisms: one shifts the voltage dependence to the negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium by charge effect, the second mechanism is less known. By applying the single-channel recordings and mutagenesis on the putative binding sites (most of them related to the selectivity filter), they concluded that the selectivity filter is stabilized to a conductive state by PUFA binding.
Strengths:
They mainly used single-channel recordings and directly assessed the behavior of the selectivity filter. The method is straightforward and convincing enough to support their claims.
Weaknesses:
The structural model they used is the KCNQ1 channel without KCNE1 because KCNQ1/KCNE1 channel complex is not available yet. As the binding site of PUFAs might overlap with KCNE1, it is not very clear how PUFA binds to the KCNQ1 channel in the presence of KCNE1.
Using other previous PUFA-related KCNQ1 mutants will strengthen their conclusions. For example, the Gmax of the K326E mutant is reduced by PUFA binding. Examining whether K326E shows reduced numbers of non-empty sweeps in the single-channel recordings will be a good addition.
We thank the reviewer for the public review. We would like to address the main weak points of the comments. As a structure of KCNQ1/KCNE1 in complex is not available yet, we used KCNQ1 alone. We believe that the PUFA and KCNE1 binding sites will not overlap as we previously presented data in agreement with the idea that KCNE1 rotates the VSD relative the PD (Wu et al., 2021). This would leave enough space for both PUFA and KCNE1, so that PUFA can bind to the crevice (K326 and D301) without competing with KCNE1. We appreciate the suggestion of adding single-channel recordings of K326E mutant and we agree it would make a valuable addition to strengthen our conclusions. However, single channel recordings for KCNQ1 are very challenging and time consuming to obtain, so we would like to keep this in consideration for future studies.
Reviewer #3 (Public Review):
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating such that the open state of the pore is unstable and undergoes intermittent closed and open conformations. PUFA enhances the maximum open probability of IKs by binding to a crevice adjacent to the pore and stabilizing the open conformation. This mechanism is supported by convincing single-channel recordings that show empty and open channel traces and the ratio of such traces is affected by PUFA. In addition, mutations of the pore residues alter PUFA effects, convincingly supporting that PUFA alters the interactions among these pore residues.
Strengths:
The data are of high quality and the description is clear.
Weaknesses:
Some comments about the presentation.
(1) The structural illustrations in this manuscript in general need to be more clarified.
(2) The manuscript heavily relies on the comparison between the S4-down and S4-up structures (Figures 3, 4, and 7) to illustrate the difference between the extracellular side of the pore and to lead to the hypothesis of open-state stability being affected by PUFA. This may mislead the readers to think that the closed conformation of the channel in the up-state is the same as that in the down-state.
We thank the reviewer for the public review, and we would like to address the comments about the presentation. We agree that the structural illustrations need to be more detailed, and we amended our previous illustrations. We have now included a new Figure 3 with a more detailed legend and a new Figure 4 that includes more information, such as the main chain of the whole selectivity filter and surrounding peptide.
We have now added some clarification regarding the structures of KCNQ1 with S4-down and S4-up to clarify that the closed conformation of the channel in the up-state is different from that in the down-state. We also emphasize this difference in the Discussion.
Recommendations for the authors:
Reviewer #1:
(1) Explain more thoroughly how the single-channel recordings were done:
- How was Lin-Glycine applied in these experiments? The patch configuration is unclear. Was Lin-Glycine added to the patch pipette? If not, why is Lin-Glycine expected to reach the proposed binding site in the outer leaflet? Were controls time-matched applications of vehicles with ethanol?
Data were collected using the cell attached patch configuration to minimize disruption to the patch and avoid rundown problems due to the loss of PIP2. Lin-Glycine was solubilized in DMSO and the desired concentration was added directly to the bath. We had no a priori reason to know if the PUFA would reach the proposed binding site but the consistency at which there was an increase in channel activity 5-10 minutes after addition to the bath convinced us that it was indeed reaching the binding site. This time frame fits with our prior experience with mefenamic acid effects on single channels (Wang et al 2020). The mefenamic acid binding site is external to the membrane so the drug must enter the cell and cross the patch membrane to affect channel activity. In addition, shown below is a previous recording from our lab, where nothing was added to the bath over a 55-minute time while recording consecutive files. This shows the typical behavior of IKs, with activity tending to cluster with a few active sweeps in between many blank sweeps. The behavior in this patch contrasts with that seen in the presence of Lin-glycine, where the clusters of activity spread over an increasing number of sweeps.
In addition, we have previously shown that 0.1% DMSO (concentration used in the present study) does not affect the GV of KCNQ1 but there is a non-significant decrease in tail current amplitudes of about 14% (Eldstrom et al., 2021). As such we do not think that the effects we see with Lin-Glycine, with an increase in activity can be explained by vehicle effects alone.
Author response image 1.
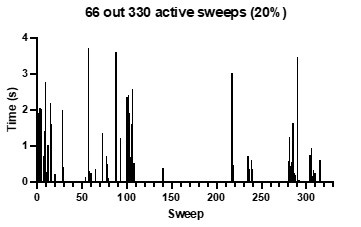
We added some more details in the section Material and Method.
- How well the replicates match the representative data in Figures 1, S1, and 6 is unclear (except for average current and Po in the last second of the traces from Figure 1). Are the results in Fig 6 n=1?
We now show in a data supplement that 3 replicates were used to access the change in channel activity upon addition of Lin-glycine.
- Diary plots (as in Werry et al. 2013) and additional descriptions of the timeline of Lin-Glycine application and analyses could add credibility to interpretations.
We added a Diary plot of for the First latency to open in Supplementary Figure S1.
- Amounts of plasmids and lipofectamine that were used in transfections are missing.
We added the information in Material and Method section as follow:
“Single channel currents were recorded from transiently transfected mouse ltk- fibroblast cells (LM cells) using 1.5 mL Lipofectamine 2000 (Thermo Fisher Scientific). Cells were transfected with 1.5 mg of pcDNA3 containing a linked KCNE1-KCNQ1 construct 20, to ensure fully KCNE1-saturated complexes, in addition to a plasmid containing green fluorescent protein (GFP) to identify transfected cells”
- Inclusion/exclusion criteria for patches analyzed are missing.
We added the information in Material and Method section as follow:
“Only patches that were largely free of endogenous currents and had few channels, such that there were several blank sweeps to average for use for leak subtraction, were analyzed.”
- Whether blinding, randomization, or pre-determined n values were employed is not mentioned.
No blinding, randomization or pre-determined n values were employed.
- Analysis methods are sometimes unclear: How was Po calculated? Representative sweeps appear to have been leak and capacitance subtracted. How was that done?
Po was estimated from all-point amplitude histogram as follow: Po = Sum (iN/(iestimateNtotal), where N is the number of points for a specific current i in the histogram, iestimate = 0.4 pA from the peak of the histogram, and Ntotal = 10,000 is the total number of points in the last second of the trace. p = 0.75 ± 0.12 (n = 8) and p = 0.87 ± 0.04 (n = 3) for Control and Lin-Glycine, respectively.
Leak and capacitance were subtracted with averaged empty sweeps.
(2) The change of cells used for whole cell vs single channel (oocytes vs mouse ltk- fibroblast cells) could be discussed. These cells likely have different lipids in their membranes. Is there any other evidence that PUFAs have the same effects on KCNE1-KCNQ1 in these cells? Does the V0.5 shift?
A similar effect on Gmax, in both oocytes and mouse ltk-fibroblast cells, is shown in Figure 1 and 2. In Figure 2, the shift in latency suggests a shift in V0.5, suggesting the binding of PUFA to Site I.
(3) The manuscript associates selectivity filter changes with S4 being up or down. It would help to clarify whether there was a change in [K+] in the two KCNQ1 structures used for modeling, as Mandala and MacKinnon (2023) state: "We note that one interesting difference between the two up structures regards the occupancy of K+ ions in the selectivity filter (SI Appendix, Fig. S5 C and D). In the polarized sample, due to the low extravesicular concentration of K+, density is only visible at the first and third positions in the selectivity filter, while density is present at all four positions in the unpolarized sample. Similar differences were observed in our previous study on Eag (20) and are qualitatively consistent with crystal structures of KcsA solved under symmetrical high and low K+ concentrations (45)."
Our studies states that there are some differences in the two structures with S4 in up-state and S4 in down-state and a reorganization of the pore. As for the change in [K+] occupancy in the two structures, we are not sure as our knowledge only come from what stated in Mandala and Mackinnon (2023). Mandala and MacKinnon did not discuss the selectivity filter in the down state structure in their paper and there are no K ions in any of their pdb files. So, we don’t know how many K+ ions there are in the down state.
(4) The manuscript states " PUFAs increase Gmax by binding to a crevice in the pore domain" and "we elucidated that Lin-Glycine binds to a crevice between K326 and D301", this seems speculative without any actual binding studies or concrete structural evidence. A quantitative structural modeling analysis of whether changes in the crevice change the theoretical binding of Lin-Glycine might provide a stronger basis for speculation.
We toned down these statements in Results and Discussion to:
“Crevice residues affect PUFA ability to increase Gmax"
And
Discussion: “We tested the hypothesis that the effect of Lin-Glycine involved conformational changes in the selectivity filter following PUFA binding to two residues K326 and D301 at the pore domain. Those residues delimit a small crevice that seems to change in size in different structures with S4 up or S4 down (Figure 3, D-F).”
(5) The several figures detailing differences in selectivity filter conformation in the KCNQ1 structures are interesting and relevant in that they identify the movement of residues such as Y315 that, when mutated, ablate Lin-Glycine effect on Gmax. It would help to clarify whether T312 and I313 also move between the two selectivity filter conformations.
From the morph of the selectivity filter in the two conformations, it is noticeable that the changes and residue movements involve only residues at the upper part of the selectivity filter (including Y315 and D317). T312 and I313, are in the lower part of the selectivity filter and do not seem to move or rotate from their position between the two conformations of the selectivity filter.
We now include a Supplementary Figures S3 and S4 that show the extent of movement of each residue in the pore region and a short description of this in the Results section.
(6) The claim in the abstract that selectivity filter instability "explains the low open probability" seems too general. Lin-Glycine seems to increase the likelihood of conduction by 2.5-fold, but it was not clear whether open probability ceases to be low or whether other mechanisms also keep Po low.
We reword this sentence to “Our results suggest that the selectivity filter in KCNQ1 is normally unstable, contributing to the low open probability, and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state..”
Reviewer #2:
(1) While all the electrophysiological recordings used KCNQ1/KCNE1 channels, all the structural models they used are KCNQ1 channels (without KCNE1). I know it is because the KCNQ1/KCNE1 complex structure is unavailable. However, according to their previous results, KCNQ1 alone is also upregulated by PUFAs. I am curious about what the single-channel recordings of KCNQ1 alone look like in the presence and absence of PUFAs.
We would love to include single-channel recordings of KCNQ1, but they are extremely hard to measure due to the small size and flickering nature of the channel.
(2) As mentioned above, we do not have the KCNQ1/KCNE1 structure yet have the KCNQ1/KCNE3 structures (Sun and MacKinnon, Cell, 2020). According to the PDBs (6V00 or 6V01), the clevis (K326 and D301) looks covered by KCNE3. Is it true that PUFAs do not upregulate KCNQ1/KCNE3? If true, KCNE1 may not cover the clevis, so the binding mode should differ from the KCNQ1/KCNE3 structures. Please discuss the possible blocking of the clevis by KCNE proteins.
We previously presented data that is consistent with that KCNE1 rotates the VSD towards the PD (Wu et al., 2021). This mechanism would leave room for PUFA and KCNE1, so that PUFA can bind to the crevice (K326 and D301). So we think that this rotation will prevent PUFA and KCNE1 from competing for the same space. As for KCNQ1/KCNE3 we currently do not have any evidence about a possible upregulation by PUFA.
(3) In the cryoEM structure with S4 resting (Figure 3F), the clevis looks too narrow for PUFA to bind. Is there any (either previous or current) evidence supporting that PUFA binding is state-dependent?
Because PUFAs integrate first into the bilayer and then diffuse towards its binding site on the channel, it would be hard to test a state-dependence of the binding. In addition, once PUFAs are in the bilayer, the rate of binding/unbinding is quite fast (within the ns range according to our previous MD simulations), whereas opening/closing rate is very slow (100 ms-s). So, the combination of slow wash in/washout, fast binding/unbinding, and slow opening/closing would make it very difficult to test the state-dependence of the binding by using a fast perfusion or different voltage protocols.
(4) In the previous report (Liin et al. Cell Reports, 2018), K326 is the most critical site for PUFA binding. Why the K326 mutants are not included in the current study? I also would like to see the single-channel recordings of the K326E mutant, which showed a smaller Gmax. Does the PUFA application reduce the probability of non-empty traces in this mutant?
As Liin et al. reported, mutations of K326 reduce the ability of PUFA to increase the Gmax. In this work, we wanted to gain further biophysical information on the mechanism that leads to an increase in Gmax, considering the knowledge we had from work conducted in our lab previously. We therefore focused here on residues downstream of K326 that we think are important for inducing the conformational changes at the selectivity filter. We agree that single channel experiments on K326E would be very interesting but that has to be for a future study.
Minor points
(1) Liin et al. used S209F (Po of 0.4) and I204F (Po of 0.04) mutants. Their single-channel recordings would be a good addition.
We thank the reviewer for the suggestion. However, single channels analysis on S209F and I204F were previously shown (Eldstrom et al., 2010).
(2) I would like to see how the Site I mutations (R2Q/Q3R) affect (or do not affect) the single-channel recordings (open probability and latency).
Thank you for the excellent suggestion. It would be interesting to assess the behavior of the channel when mutations occur at Site I. However, we think this information will not add any more detail to this study as we focus here our attention on the mechanism for Gmax increase. Single channels recordings are extremely hard to get, therefore we chose to include only mutations at Site II for this study.
(3) I would like the G-V curves for all the mutations at 0 and 20 uM of Lin-Glycine (Figure 3C and Figures 5A and B).
We now added the G-V curves in Supplementary Figure S7.
(4) I assume all the PUFAs have a similar effect on the selectivity filter, but a few other examples of PUFAs would be nice to see.
We anticipate that PUFAs and analogues with similar properties to Lin-Glycine would increasing the Gmax by a similar mechanism, because other PUFAs have been previously shown to increase the Gmax (Bohannon et al., 2020).
(5) Although the probabilities of non-empty sweeps are written in the manuscript, bar graph presentations would be a nice addition to Figures 2 and 6.
We have added bar graphs of non-empty sweeps for Fig 2 and 6 in.
(6) Is there no statistical significance for D317E and T309S in Figure 5A?
No statistical significance for D317E and T309S
(7) There is no reference to Figure 7 in the manuscript.
A reference to Figure 7 has been added to the manuscript in the following paragraph.
“Taken together, our results suggest that the binding of PUFA to Site II increases Gmax by promoting a series of interactions that stabilize the channel pore in the conductive state. For instance, we speculate that in the conductive state, hydrogen bonds between W304-D317 and W305-Y315, which are likely absent in the non-conductive conformation of KCNQ1, are created and that PUFA binding to Site II favors the transition towards the conductive state of the channel (Figure 7)”
Reviewer #3:
(1) Clarify the structural figures. Figures 3 D, E, and F - explain what the colors indicate.
A more detailed description of Figure 3 has been added to the legend.
“D, E and F) Structure of crevice between S5 and S6 in KCNQ1 with S4 up (D and E) and S4 down (F). Residues that surround the crevice from S6 shown in blue (K326, T327, S330, V334) and from S5 in red (D301, A300, L303, F270). Remaining KCNQ1 residues shown in purple…, linoleic acid (LIN: gold color)”
Fig 4. Only side chains of the residues are shown, making it hard to relate the figure to the familiar K channel selectivity filter. The main chain of the entire selectivity should be shown to orient readers to the familiar view of the K channel selectivity filter. In addition, the structures shown are only part of the selectivity filter, it should be specified which part of the selectivity filter is shown. These will also help the discussion at the bottom of page 10 and subsequent text.
We now provide a new Figure 4 with more details such as the main chain of the whole selectivity filter and surrounding peptide.
(2) Cautions should be stated clearly when the structural comparison between the S4-up and S4-down is made that the structure of the pore when it is closed with S4-up may differ from the structure of the pore with S4-down.
We now state in addition “Clearly, there will be other differences in the pore domain between structures with activated and resting VSDs, for example the state of the activation gate.”
-
eLife assessment
This study reveals an important mechanism, a polyunsaturated fatty acid increases a K+ channel conductance by helping its K+ selectivity filter form a conductive state. Overall, this mechanism is supported by convincing single channel recordings, macroscopic current recordings and mutational analyses, though further clarification of some results seems warranted. These findings are expected to be of interest to researchers studying ion channel gating.
-
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single channel analysis, though some technical details are missing and presentations confusing. The linkage to PUFA action through the selectivity filter is supported by disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. A definitive functional experiment is conducted by single channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. The computational exploration of two selectivity filter structures proposed to …
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single channel analysis, though some technical details are missing and presentations confusing. The linkage to PUFA action through the selectivity filter is supported by disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. A definitive functional experiment is conducted by single channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. The computational exploration of two selectivity filter structures proposed to interact distinctly with Lin-Glycine is informative, however the relation of the closed selectivity filter structures to the [K+] concentration in which it was obtained and inactivation in other channels is ignored. Overall, the major claim of the abstract is well-supported: "... that the selectivity filter in KCNQ1 is normally unstable ... and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state."
-
Reviewer #2 (Public Review):
Summary:
Golluscio et al. addresses one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by a polyunsaturated fatty acid (PUFA). PUFAs are known to upregulate KCNQ1 and KCNQ1/KCNE1 channels by two mechanisms: one shifts the voltage dependence to the negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium by charge effect, the second mechanism is less known. By applying the single-channel recordings and mutagenesis on the putative binding sites (most of them related to the selectivity filter), they concluded that the selectivity filter is stabilized to a conductive state by PUFA binding.
Strengths:
The manuscript employed single-channel recordings and directly assessed the behavior of the selectivity …
Reviewer #2 (Public Review):
Summary:
Golluscio et al. addresses one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by a polyunsaturated fatty acid (PUFA). PUFAs are known to upregulate KCNQ1 and KCNQ1/KCNE1 channels by two mechanisms: one shifts the voltage dependence to the negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium by charge effect, the second mechanism is less known. By applying the single-channel recordings and mutagenesis on the putative binding sites (most of them related to the selectivity filter), they concluded that the selectivity filter is stabilized to a conductive state by PUFA binding.
Strengths:
The manuscript employed single-channel recordings and directly assessed the behavior of the selectivity filter. The method is straightforward and convincing enough to support the claims.
Weaknesses:
Although the analysis using selectivity filter mutants supports the hypothesis that PUFA binding stabilizes the conducting state of the filter, it may be somewhat speculative how PUFAs bind to the KCNQ1 channel in the presence of KCNE1.
-
Reviewer #3 (Public Review):
Summary:
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating such that the open state of the pore is unstable and undergoes intermittent closed and open conformations. PUFA enhances the maximum open probability of IKs by binding to a crevice adjacent to the pore and stabilize the open conformation. This mechanism is supported by convincing single channel recordings that show empty and open channel traces and the ratio of such traces is affected by PUFA. In addition, mutations of the pore residues alter PUFA effects, convincingly supporting that PUFA alters the interactions among these pore residues.
Strengths:
The data are of high quality and the description is clear.
-
-
eLife assessment
This study makes a valuable finding, a polyunsaturated fatty acid increases the conductance of a K+ channel by helping its K+ selectivity filter form a conductive state. Overall, support for this major claim is solid, though other claims remain speculative with incomplete support. These findings are expected to be of interest to researchers studying ion channel gating.
-
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single-channel analysis, though some method details are missing. The linkage to PUFA action through the selectivity filter is supported by the disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. Claims about differences in Lin-Glycine binding to these two structural conformations seem to lack clear support, thus the claim seems speculative that PUFAs increase Gmax by binding to a crevice in the pore domain. A potentially definitive functional experiment is conducted by …
Reviewer #1 (Public Review):
This study makes an interesting finding: a polyunsaturated fatty acid, Lin-Glycine, increases the conductance of KCNQ1/KCNE1 channels by stabilizing a state of the selectivity filter that allows K+ conduction. The stabilization of a conducting state appears well supported by single-channel analysis, though some method details are missing. The linkage to PUFA action through the selectivity filter is supported by the disruption of PUFA effects by mutation of residues which change conformation in two KCNQ1 structures from the literature. Claims about differences in Lin-Glycine binding to these two structural conformations seem to lack clear support, thus the claim seems speculative that PUFAs increase Gmax by binding to a crevice in the pore domain. A potentially definitive functional experiment is conducted by single-channel recordings with selectivity filter domain mutation Y315F which ablates the Lin-Glycine effect on Gmax. However, this appears to be an n=1 experiment. Overall, the major claim of the abstract is supported: "... that the selectivity filter in KCNQ1 is normally unstable ... and that the PUFA-induced increase in Gmax is caused by a stabilization of the selectivity filter in an open-conductive state." However, the claim in the abstract that selectivity filter instability "explains the low open probability" seems too general.
-
Reviewer #2 (Public Review):
Summary:
Golluscio et al. address one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by polyunsaturated fatty acids (PUFA). PUFA is known to upregulate KCNQ1 and KCNQ1/KCNE1 channels by two mechanisms: one shifts the voltage dependence to the negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium by charge effect, the second mechanism is less known. By applying the single-channel recordings and mutagenesis on the putative binding sites (most of them related to the selectivity filter), they concluded that the selectivity filter is stabilized to a conductive state by PUFA binding.
Strengths:
They mainly used single-channel recordings and directly assessed the behavior of the selectivity filter. The …Reviewer #2 (Public Review):
Summary:
Golluscio et al. address one of the mechanisms of IKs (KCNQ1/KCNE1) channel upregulation by polyunsaturated fatty acids (PUFA). PUFA is known to upregulate KCNQ1 and KCNQ1/KCNE1 channels by two mechanisms: one shifts the voltage dependence to the negative direction, and the other increases the maximum conductance (Gmax). While the first mechanism is known to affect the voltage sensor equilibrium by charge effect, the second mechanism is less known. By applying the single-channel recordings and mutagenesis on the putative binding sites (most of them related to the selectivity filter), they concluded that the selectivity filter is stabilized to a conductive state by PUFA binding.
Strengths:
They mainly used single-channel recordings and directly assessed the behavior of the selectivity filter. The method is straightforward and convincing enough to support their claims.Weaknesses:
The structural model they used is the KCNQ1 channel without KCNE1 because KCNQ1/KCNE1 channel complex is not available yet. As the binding site of PUFAs might overlap with KCNE1, it is not very clear how PUFA binds to the KCNQ1 channel in the presence of KCNE1.Using other previous PUFA-related KCNQ1 mutants will strengthen their conclusions. For example, the Gmax of the K326E mutant is reduced by PUFA binding. Examining whether K326E shows reduced numbers of non-empty sweeps in the single-channel recordings will be a good addition.
-
Reviewer #3 (Public Review):
Summary:
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating such that the open state of the pore is unstable and undergoes intermittent closed and open conformations. PUFA enhances the maximum open probability of IKs by binding to a crevice adjacent to the pore and stabilizing the open conformation. This mechanism is supported by convincing single-channel recordings that show empty and open channel traces and the ratio of such traces is affected by PUFA. In addition, mutations of the pore residues alter PUFA effects, convincingly supporting that PUFA alters the interactions among these pore residues.
Strengths:
The data are of high quality and the description is clear.Weaknesses:
Some comments about the presentation.(1) The structural illustrations in this manuscript in general need …
Reviewer #3 (Public Review):
Summary:
This manuscript reveals an important mechanism of KCNQ1/IKs channel gating such that the open state of the pore is unstable and undergoes intermittent closed and open conformations. PUFA enhances the maximum open probability of IKs by binding to a crevice adjacent to the pore and stabilizing the open conformation. This mechanism is supported by convincing single-channel recordings that show empty and open channel traces and the ratio of such traces is affected by PUFA. In addition, mutations of the pore residues alter PUFA effects, convincingly supporting that PUFA alters the interactions among these pore residues.
Strengths:
The data are of high quality and the description is clear.Weaknesses:
Some comments about the presentation.(1) The structural illustrations in this manuscript in general need to be more clarified.
(2) The manuscript heavily relies on the comparison between the S4-down and S4-up structures (Figures 3, 4, and 7) to illustrate the difference between the extracellular side of the pore and to lead to the hypothesis of open-state stability being affected by PUFA. This may mislead the readers to think that the closed conformation of the channel in the up-state is the same as that in the down-state.
-
