Interaction hierarchy among Cdv proteins drives recruitment to membrane necks
Curation statements for this article:-
Curated by eLife

eLife Assessment
This important study investigates how the Cdv division proteins of Metallosphaera sedula assemble on and interact with curved membranes in vitro, advancing our understanding of this reduced ESCRT-like machinery. The data provide support for sequential protein recruitment and curvature-dependent enrichment at membrane necks, based on well-controlled reconstitution assays and quantitative analysis. The work establishes a convincing experimental framework for dissecting Cdv-mediated membrane remodeling. The study will be of broad interest to evolutionary and synthetic biologists as well as membrane biophysicists.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Cell division in the crenarchaea is accomplished by the Cdv system. In Sulfolobus cells, it was observed that an initial non-contractile ring of CdvA and CdvB forms at the mid location of the cell, which is followed by a second ring of CdvB1 and CdvB2 that appear to drive the constriction of the cell membrane. Here, we use an in vitro reconstituted system to explore how protein interactions among these Cdv proteins govern their recruitment to the membrane. We show that CdvA can bind to lipid membranes, but does so more efficiently when it is in complex with CdvB. We find that CdvB2 can polymerize if its self-inhibitory domain is removed, and that by itself is exhibits poor binding to the membrane. However, CdvB2 can be efficiently recruited to the membrane by both CdvB1 and CdvB. Furthermore, the CdvB1:CdvB2 co-polymer can be recruited to the membrane by CdvA:CdvB. By reconstituting these proteins in dumbbell-shaped liposomes, we show that Cdv proteins have a strong preference to localize at membrane necks of high curvature. Our findings clarify many of the mutual protein interactions of the Cdv system and their interaction with the membrane, thus helping to build a mechanistic understanding of cell division in archaeal cells.
Article activity feed
-

-
-

eLife Assessment
This important study investigates how the Cdv division proteins of Metallosphaera sedula assemble on and interact with curved membranes in vitro, advancing our understanding of this reduced ESCRT-like machinery. The data provide support for sequential protein recruitment and curvature-dependent enrichment at membrane necks, based on well-controlled reconstitution assays and quantitative analysis. The work establishes a convincing experimental framework for dissecting Cdv-mediated membrane remodeling. The study will be of broad interest to evolutionary and synthetic biologists as well as membrane biophysicists.
-
Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbell-shaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two sub-complexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane necks. Similarly, the findings on CdvB1:CdvB2ΔC indicate that truncation of CdvB2 facilitates …Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbell-shaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two sub-complexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane necks. Similarly, the findings on CdvB1:CdvB2ΔC indicate that truncation of CdvB2 facilitates filament formation and enhances curvature sensitivity in interaction with CdvB1. Finally, the authors reconstitute a quaternary CdvA:CdvB:CdvB1:CdvB2 complex and demonstrate its enrichment at membrane necks. The mechanistic details of how these complexes drive membrane remodeling, particularly through subcomplex removal by the proteasome and/or CdvC, remain insufficiently addressed, and the study therefore mainly provides an experimental framework for future mechanistic investigation.Strengths:
The study of machinery assembly and its involvement in membrane remodeling, particularly using bottom-up reconstituted in vitro systems, presents significant challenges. This is particularly true for systems like the ESCRT-III complex, which localizes uniquely at the lumen of membrane necks prior to scission. The use of dumbbell-shaped liposomes in this study provides a promising experimental model to investigate ESCRT-III and ESCRT-III-like protein activity at membrane necks.
The authors present intriguing evidence regarding the sequential recruitment of ESCRT-III proteins in crenarchaea-a close relative of eukaryotes.Weaknesses:
The findings of this study suggest that the hierarchical recruitment characteristic of eukaryotic systems may predate eukaryogenesis, which represents a significant and exciting contribution. However, the broader implications of these findings for membrane remodeling mechanisms remain largely unexplored. Nevertheless, this study provides a valuable experimental framework to address these questions in the future.
-
Reviewer #2 (Public review):
Summary:
The Crenarchaeal Cdv division system represents a reduced form of the universal and ubiquitous ESCRT membrane reverse-topology scission machinery, and therefore a prime candidate for synthetic and reconstitution studies. The work here represents a convincing extension of previous work in the field, clarifying the order of recruitment of Cdv proteins to curved membranes.
Strengths:
The use of a recently developed approach to produce dumbbell-shaped liposomes (De Franceschi et al. 2022), which allowed the authors to assess recruitment of various Cdv assemblies to curved membranes or membrane necks; reconstitution of a quaternary Cdv complex at a membrane neck.
Weaknesses:
The initial manuscript was a bit light on quantitative detail, across the various figures - addressing this would make the paper …
Reviewer #2 (Public review):
Summary:
The Crenarchaeal Cdv division system represents a reduced form of the universal and ubiquitous ESCRT membrane reverse-topology scission machinery, and therefore a prime candidate for synthetic and reconstitution studies. The work here represents a convincing extension of previous work in the field, clarifying the order of recruitment of Cdv proteins to curved membranes.
Strengths:
The use of a recently developed approach to produce dumbbell-shaped liposomes (De Franceschi et al. 2022), which allowed the authors to assess recruitment of various Cdv assemblies to curved membranes or membrane necks; reconstitution of a quaternary Cdv complex at a membrane neck.
Weaknesses:
The initial manuscript was a bit light on quantitative detail, across the various figures - addressing this would make the paper much stronger. The authors could also include in the discussion a short paragraph on implications for our understanding of ESCRT function in other contexts and/or in archaeal evolution - for the interests of a broad audience. These issues have been addressed in the authors' revision.
-
Reviewer #3 (Public review):
In this revised report, De Franceschi et al. purify components of the Cdv machinery in archaeon M. sedula and probe their interactions with membrane and with one-another in vitro using two main assays - liposome flotation and fluorescent imaging of encapsulated proteins. This has the potential to add to the field by showing how the order of protein recruitment seen in cells is related to the differential capacity of individual proteins to bind membranes when alone or when combined.
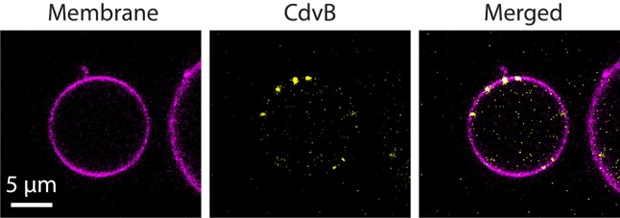
Using the floatation assay, they demonstrate that CdvA, CdvB, and CdvB1 bind liposomes. CdvB2 lacking its C-terminus is not efficiently recruited to membranes unless CdvAB or CdvB1 are present. The authors then employ a clever liposome assay that generates chained spherical liposomes connected by thin membrane necks, which allows …
Reviewer #3 (Public review):
In this revised report, De Franceschi et al. purify components of the Cdv machinery in archaeon M. sedula and probe their interactions with membrane and with one-another in vitro using two main assays - liposome flotation and fluorescent imaging of encapsulated proteins. This has the potential to add to the field by showing how the order of protein recruitment seen in cells is related to the differential capacity of individual proteins to bind membranes when alone or when combined.
Using the floatation assay, they demonstrate that CdvA, CdvB, and CdvB1 bind liposomes. CdvB2 lacking its C-terminus is not efficiently recruited to membranes unless CdvAB or CdvB1 are present. The authors then employ a clever liposome assay that generates chained spherical liposomes connected by thin membrane necks, which allows them to accurately control the buffer composition inside and outside of the liposome. With this, they show that all four proteins accumulate in necks of dumbbell-shaped liposomes that mimic the shape of constricting necks in cell division, possibly indicating a sensing of catenoid membrane geometry. Taken altogether, these data lead them to propose that Cdv proteins are sequentially recruited to the membrane as has also been suggested by in vivo studies of ESCRT-III dependent cell division in crenarchaea.
In their revision, the authors have addressed the vast majority of our previous concerns. The paper is much improved as a result. The Figures are improved and the authors have added appropriate controls and additional experiments, strengthening their conclusions.
There are still some discrepancies between these results and what is know about Sulfolobus division. Since the initial submission, other work has shown that in S. acidocaldarius, CdvA is the first component to assemble a ring (in absence of CdvB , doi.org/10.1073/pnas.2513939122) and that CdvB2 is able to bind membranes in vitro (doi.org/10.1073/pnas.2525941123). This might reflect differences between Sulfolobus and Metallosphaera, but probably should be discussed.
-
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbellshaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two subcomplexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane …
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbellshaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two subcomplexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane necks. Similarly, the findings on CdvB1:CdvB2ΔC indicate that truncation of CdvB2 facilitates filament formation and enhances curvature sensitivity in interaction with CdvB1. Finally, while the authors reconstitute a quaternary CdvA:CdvB:CdvB1:CdvB2 complex and demonstrate its enrichment at membrane necks, the mechanistic details of how these complexes drive membrane remodeling by subcomplexes removal by the proteasome and/or CdvC remain speculative.
Although the work highlights intriguing similarities with eukaryotic ESCRT-III systems and explores unique archaeal adaptations, the conclusions drawn would benefit from stronger experimental validation and a more comprehensive mechanistic framework.
Strengths:
The study of machinery assembly and its involvement in membrane remodeling, particularly using bottom-up reconstituted in vitro systems, presents significant challenges. This is particularly true for systems like the ESCRT-III complex, which localizes uniquely at the lumen of membrane necks prior to scission. The use of dumbbell-shaped liposomes in this study provides a promising experimental model to investigate ESCRT-III and ESCRT-III-like protein activity at membrane necks.
The authors present intriguing evidence regarding the sequential recruitment of ESCRT-III proteins in crenarchaea-a close relative of eukaryotes. This finding suggests that the hierarchical recruitment characteristic of eukaryotic systems may predate eukaryogenesis, which is a significant and exciting contribution. However, the broader implications of these findings for membrane remodeling mechanisms remain speculative, and the study would benefit from stronger experimental validation and expanded contextualization within the field.
We thank the Referee for his/her appreciation of our work.
Weaknesses:
This manuscript presents several methodological inconsistencies and lacks key controls to validate its claims. Additionally, there is insufficient information about the number of experimental repetitions, statistical analyses, and a broader discussion of the major findings in the context of open questions in the field.
We have now added more controls, information about repetitions, and discussion.
Reviewer #2 (Public review):
Summary:
The Crenarchaeal Cdv division system represents a reduced form of the universal and ubiquitous ESCRT membrane reverse-topology scission machinery, and therefore a prime candidate for synthetic and reconstitution studies. The work here represents a solid extension of previous work in the field, clarifying the order of recruitment of Cdv proteins to curved membranes.
Strengths:
The use of a recently developed approach to produce dumbbell-shaped liposomes (De Franceschi et al. 2022), which allowed the authors to assess recruitment of various Cdv assemblies to curved membranes or membrane necks; reconstitution of a quaternary Cdv complex at a membrane neck.
We thank the Referee for his/her appreciation of the work.
Weaknesses:
The manuscript is a bit light on quantitative detail, across the various figures, and several key controls are missing (CdvA, B alone to better interpret the co-polymerisation phenotypes and establish the true order of recruitment, for example) - addressing this would make the paper much stronger. The authors could also include in the discussion a short paragraph on implications for our understanding of ESCRT function in other contexts and/or in archaeal evolution, as well as a brief exploration of the possible reasons for the discrepancy between the foci observed in their liposome assays and the large rings observed in cells - to better serve the interests of a broad audience.
We have now added more controls, information about repetitions, and discussion.
Reviewer #3 (Public review):
Summary:
In this report, De Franceschi et al. purify components of the Cdv machinery in archaeon M. sedula and probe their interactions with membrane and with one-another in vitro using two main assays - liposome flotation and fluorescent imaging of encapsulated proteins. This has the potential to add to the field by showing how the order of protein recruitment seen in cells is related to the differential capacity of individual proteins to bind membranes when alone or when combined.
Strengths:
Using the floatation assay, they demonstrate that CdvA and CdvB bind liposomes when combined. While CdvB1 also binds liposomes under these conditions, in the floatation assay, CdvB2 lacking its C-terminus is not efficiently recruited to membranes unless CdvAB or CdvB1 are present. The authors then employ a clever liposome assay that generates chained spherical liposomes connected by thin membrane necks, which allows them to accurately control the buffer composition inside and outside of the liposome. With this, they show that all four proteins accumulate in necks of dumbbell-shaped liposomes that mimic the shape of constricting necks in cell division. Taken altogether, these data lead them to propose that Cdv proteins are sequentially recruited to the membrane as has also been suggested by in vivo studies of ESCRT-III dependent cell division in crenarchaea.
We thank the Referee for his/her appreciation of the work.
Weaknesses:
These experiments provide a good starting point for the in vitro study the interaction of Cdv system components with the membrane and their consecutive recruitment. However, several experimental controls are missing that complicate their ability to draw strong conclusions. Moreover, some results are inconsistent across the two main assays which make the findings difficult to interpret:
(1) Missing controls.
Various protein mixtures are assessed for their membrane-binding properties in different ways. However, it is difficult to interpret the effect of any specific protein combination, when the same experiment is not presented in a way that includes separate tests for all individual components. In this sense, the paper lacks important controls. For example, Fig 1C is missing the CdvB-only control. The authors remark that CdvB did not polymerise (data not shown) but do not comment on whether it binds membrane in their assays. In the introduction, Samson et al., 2011 is cited as a reference to show that CdvB does not bind membrane. However, here the authors are working with protein from a different organism in a different buffer, using a different membrane composition and a different assay. Given that so many variables are changing, it would be good to present how M. sedula CdvB behaves under these conditions.
We thank the referee for raising this point. We have now added these data in Figure 1C. Indeed it turns out that CdvB from M. sedula exhibits clear membrane binding on its own in a flotation assay.
Similarly, there is no data showing how CdvB alone or CdvA alone behave in the dumbbell liposome assay.
Without these controls, it's impossible to say whether CdvA recruits CdvB or the other way around. The manuscript would be much stronger if such data could be added.
We have now added these data in Figure 1E, 1F and 1G. Overall, we can confirm that CdvA binds the membrane better in the presence of CdvB (although both proteins can bind the membrane on their own). Both proteins appear to recognize the curved region of the membrane neck.
(2) Some of the discrepancies in the data generated using different assays are not discussed.
The authors show that CdvB2∆C binds membrane and localizes to membrane necks in the dumbbell liposome assay, but no membrane binding is detected in the flotation assay. The discrepancy between these results further highlights the need for CdvB-only and CdvA-only controls.
We have now added these controls in Figure 1. In addition, we would like to clarify that the flotation assay and the SMS dumbbell assay serve different purposes and are not directly comparable in quantitative terms. In the flotation assay, all the protein present as input is eventually recovered and visualized. Thus, quantitative information on the proportion of the fraction of the total protein bound to lipids can be inferred from this assay. The SMS assay, in contrast, provides a very different kind of information. Because of the particular protocol required to generate dumbbells (De Franceschi, 2022), the total amount of protein in the inner buffer in dumbbells is not accurately defined, because protein that is not correctly reconstituted (e.g. which aggregates while still in the droplet phase) will interfere with vesicle generation, with the result that dumbbell with such aggregates is generally not formed in the first place. This renders it impossible to draw any quantitative conclusions about the proportion of the sample bound to lipids. The SMS is therefore not directly comparable to the flotation assay, and it is rather complementary to it. Indeed, the purpose of the SMS is to provide information about curvature selectivity of the protein.
(3) Validation of the liposome assay.
The experimental setup to create dumbbell-shaped liposomes seems great and is a clever novel approach pioneered by the team. Not only can the authors manipulate liposome shape, they also state that this allows them to accurately control the species present on the inside and outside of the liposome. Interpreting the results of the liposome assay, however, depends on the geometry being correct. To make this clearer, it would seem important to include controls to prove that all the protein imaged at membrane necks lie on the inside of liposomes. In the images in SFig3 there appears to be protein outside of the liposome. It would also be helpful to present data to show test whether the necks are open, as suggested in the paper, by using FRAP or some other related technique.
We thank the Referee for his/her appreciation. The proteins are encapsulated inside the liposomes, not outside of them. While Figure S3 might give the appearance that there is some protein outside, this is actually just an imaging artifact. Author response image 1 (below) explains this: When the membrane and protein channel are shown separately, it is clear that the protein cluster that appeared to be ‘outside’ actually colocalizes with an extra small dumbbell lobe (yellow arrowhead). The protein appeared to be outside of it because (1) the protein fluorescent signal is stronger than the signal from the membrane, and (2) there is a certain time delay in the acquisition of the two channels (0.5-1 second), thus the membrane may have slightly shifted out of focus when the fluorescence was being acquired. We are confident that the protein is inside in these dumbbells because the procedure for preparing the dumbbells requires extensive emulsification by pipetting, which requires ≈ 1 minute. This time is more than sufficient for proteins with high affinity for the membrane, like ESCRT and Cdv, to bind the membrane. For an example of how fast binding under confinement can be, please see movie 2 from this paper: De Franceschi N, Alqabandi M, Miguet N, Caillat C, Mangenot S, Weissenhorn W, Bassereau P. The ESCRT protein CHMP2B acts as a diffusion barrier on reconstituted membrane necks. J Cell Sci. 2018 Aug 3;132(4):jcs217968.
Moreover, in many instances, we observed that the protein is inside because, by increasing the gain in the images post-acquisition, a clear protein signal appear in the lumen (see Author response image 2).
Author response image 1.
Separate channels showing colocalization of protein and lipids (adapted from Figure S3). The zoom-in shows separate channels, highlighting that the CdvB2 cluster that seems to be ‘outside the dumbbell’ actually colocalizes with the small terminal lobe of the dumbbell, indicating that the protein is encapsulated within that lobe.
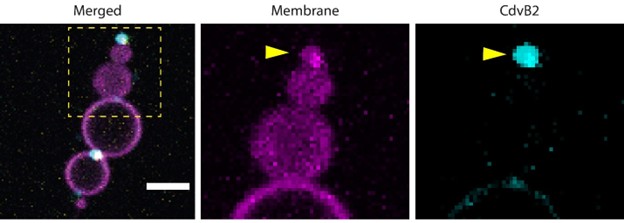
Author response image 2.
Residual protein present inside lumen of dumbbells as visualized by increasing the brightness post-acquisition.
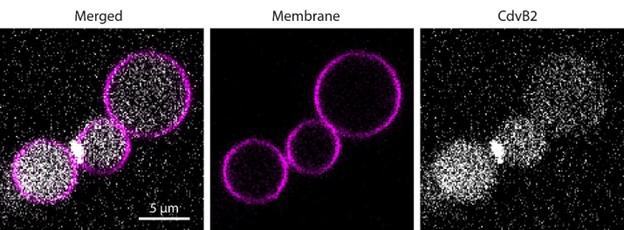
We are not sure what the referee means by “test whether the necks are open, as suggested in the paper”. We are confident that the lobes of dumbbells originated from a single floppy vesicle, and were therefore mutually connected with an open neck (at least at the onset of the experiment). We have performed extensive FRAP assays on dumbbells in previous papers (De Franceschi et al., ACS nano 2022 and De Franceschi et al., Nature Nanotech 2024) which unequivocally proved that these chains of dumbbells are connected with open necks. We now also performed a few FRAP assay with reconstituted Cdv proteins, which confirmed this point. We have added a movie of such an experiment to the manuscript (Movie 1).
Investigating whether the necks are open or closed after Cdv reconstitution is indeed a very relevant question, that could be rephrased as “verify whether Cdv proteins or their combination can induce membrane scission”. This is however beyond the scope of this manuscript, as the current work merely addressed the question of hierarchical recruitment of Cdv proteins at the membrane. We plan to examine this in future work.
(4) Quantification of results from the liposome assay.
The paper would be strengthened by the inclusion of more quantitative data relating to the liposome assay. Firstly, only a single field of view is shown for each condition. Because of this, the reader cannot know whether this is a representative image, or an outlier? Can the authors do some quantification of the data to demonstrate this? The line scan profiles in the supplemental figures would be an example of this, but again in these Figures only a single image is analyzed.
The images that we showed are indeed representative. The dumbbells that are generated by the SMS approach contain an “internal control”: in each dumbbell, the protein has the option of localizing at the neck or localizing elsewhere in the region of flat membrane. We see consistently that Cdv proteins have a strong preference for localizing at the neck.
We would recommend that the authors present quantitative data to show the extent of co-localization at the necks in each case. They also need a metric to report instances in which protein is not seen at the neck, e.g. CdvB2 but not CdvB1 in Fig2I, which rules out a simple curvature preference for CdvB2 as stated in line 182.
While the request for better quantitation is reasonable, this would require carrying out very significant new experiments at the microscope, which is rendered near-impossible since both first authors left the lab on to new positions.
Secondly, the authors state that they see CdvB2∆C recruited to the membrane by CdvB1 (lines 184-187, Fig 2I). However, this simple conclusion is not borne out in the data. Inspecting the CdvB2∆C panels of Fig 2I, Fig3C, and Fig3D, CdvB2∆C signal can be seen at positions which don't colocalize with other proteins. The authors also observe CdvB2∆C localizing to membrane necks by itself (Fig 2E). Therefore, while CdvB1 and CdvB2∆C colocalize in the flotation assay, there is no strong evidence for CdvB2∆C recruitment by CdvB1 in dumbbells. This is further underscored by the observation that in the presented data, all Cdv proteins always appear to localize at dumbbell necks, irrespective of what other components are present inside the liposome. Although one nice control is presented (ZipA), this suggests that more work is required to be sure that the proteins are behaving properly in this assay. For example, if membrane binding surfaces of Cdv proteins are mutated, does this lead to the accumulation of proteins in the bulk of the liposome as expected?
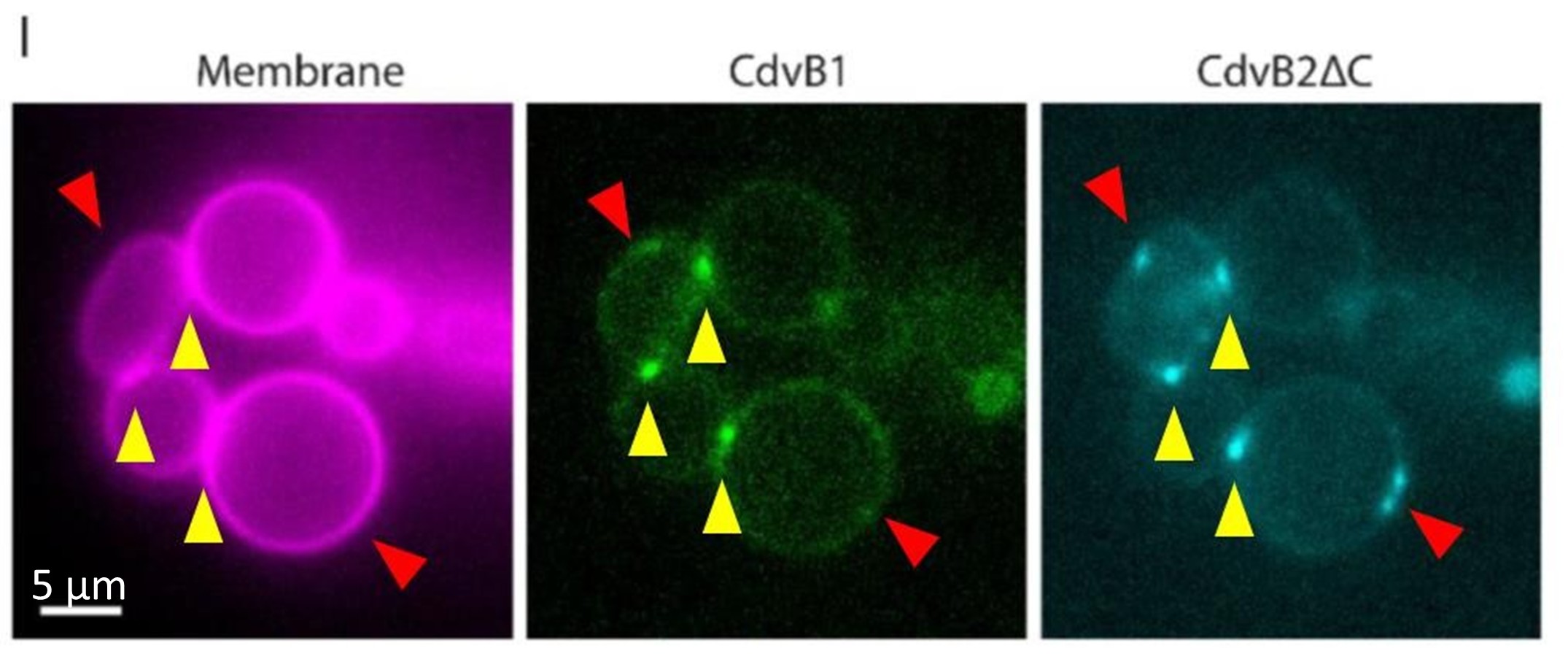
In the particular example of Figure 2I, it indeed appears that there are some clusters of CdvB2ΔC that do not contain CdvB1 (we indicated them in Author response image 3 by red arrowheads), while the yellow arrowheads indicate clusters that contain both proteins. It can be clearly seen that the clusters that do contain both proteins (yellow arrows) are localized at necks, while those that only contain CdvB2ΔC (red arrows) are not localized at necks. This is no coincidence. The clusters indicated by the red arrow do contain CdvB1. However, these clusters rapidly diffuse on the membrane plane because they are not fixed at the neck: therefore, they constantly shift in and out of focus. Because there is a time delay in the acquisition of each channel (between 0.5 and 1 second), these cluster were in focus when the CdvB2ΔC signal was being acquired, but sifted out of focus when the CdvB1 signal was being acquired. This implies that the clusters indicated by the yellow arrowheads are stably localized at necks, which is precisely the point we wished to make with this experiment: because Cdv proteins have an affinity for curved geometry, they preferentially and stably localize at necks. Why don’t all the clusters localize at necks then? We estimate that the simple answer is that, in this particular case, there are more clusters than there are necks, so some of the clusters must necessarily localize somewhere else.
Author response image 3.
Current Figure 2H, where clusters that are double-positive for both CdvB1 and CdvB2ΔC are indicated by yellow arrowheads, while cluster that apparently only contain CdvB2ΔC are indicated by red arrowheads. It is observed that all the double-positive clusters are localized at necks.
(5) Rings.
The authors should comment on why they never observe large Cdv rings in their experiments. In crenarchaeal cell division, CdvA and CdvB have been observed to form large rings in the middle of the 1 micron cell, before constriction. Only in the later stages of division are the ESCRTs localized to the constricting neck, at a time when CdvA is no longer present in the ring. Therefore, if the in vitro assay used by the authors really recapitulated the biology, one would expect to see large CdvAB rings in Figs 1EF. This is ignored in the model. In the proposed model of ring assembly (line 252), CdvAB ring formation is mentioned, but authors do not discuss the fact that they do not observe CdvAB rings - only foci at membrane necks. The discussion section would benefit from the authors commenting on this.
The referee is correct: it is intriguing that we don’t see micron-sized rings for CdvA and CdvB. We do note that our EM data (Fig.S1) show that CdvA in its own can form rings of about 100-200nm diameter, well below the diffraction limit, that could well correspond to the foci that we optically resolve in Figure 1. We now added a brief comment on this to the manuscript on lines 256-264.
(6) Stoichiometry
It is not clear why 100% of the visible CdvA and 100% of the the visible CdvB are shifted to the lipid fraction in 1C. Perhaps this is a matter of quantification. Can the authors comment on the stoichiometry here?
We agree that this was unclear. Since that particular gel was stained by coumassie, the quantitative signals might be unreliable, and hence we have repeated this experiment using fluorescently labelled proteins, which show indeed a less extreme distribution. This was also done to make the data more uniform, as requested by the referees.
(7) Significance of quantification of MBP-tagged filaments.
Authors use tagging and removal of MBP as a convenient, controllable system to trigger polymerisation of various Cdv proteins. However, it is unclear what is the value and significance of reporting the width and length of the short linear filaments that are formed by the MBP-tagged proteins. Presumably they are artefactual assemblies generated by the presence of the tag?
Providing a measure of the changes induced by MBP removal, in fact, validates that this actually has an effect. But perhaps this places too much emphasis on the short filaments. We now opted for a compromise, removing the quantification of the width and length of short filaments formed by MBPtagged protein from the text, but keeping the supplementary figure showing their distribution as compared to the other filaments (Figure S2E, SF).
Similar Figure 2C doesn't seem a useful addition to the paper.
We removed panel 2C, and now merely report these values in the text.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
I would suggest the authors perform a deeper discussion about their findings, such as what are the evolutionary implications, how they think lipids from these archaea may affect the recruitment process,...
Because there is no exact homology between Archaea Cdv proteins and Eukaryotic ESCRT-III proteins, we do not feel our work brings new evolutionary implications beyond what we already state in the manuscript. We also dis not perform experiments using Archaea lipids, thus we would rather not speculate on how they may potentially affect the recruitment of Cdv proteins.
In general, the manuscript lacks information regarding some scale bars, number of experimental repetitions (n or N), statistical analysis when needed, information about protein concentrations used in their assays.
We have now added this information in the manuscript.
Below, I provide a list of comments that I think the authors should address to improve the manuscript:
(1) Line 113-114: The authors test protein-membrane interactions using flotation assays with positively curved SUV membranes but encapsulate proteins in dumbbell-shaped liposomes with negative curvature at the connecting necks. Might the use of membranes with opposite curvatures affect the recruitment process? Since the proteins are fluorescently labeled, I suggest testing recruitment using flat giant unilamellar vesicles or supported lipid bilayers (with zero curvature) to validate their findings.
We thank the referee for this suggestion. Please do note that we are not claiming in our paper that Cdv proteins recognize negative curvature. We merely observe that they localize at necks. The neck of a dumbbell exhibits the so-called “catenoid” geometry, which is characterized by having both positive and negative curvature.
Experimentally, on the SUVs, we now realize there was a mistake in the method section: In the flotation assay we in fact used multilamellar vesicles, not SUVs, precisely for the reason mentioned by the referee. We apologize for the oversight and have now corrected this in the methods. Multilamellar vesicles are not characterized by a strong positive curvature as SUVs do, but we do agree that they likely don’t have negative curvature there either. Because of the heterogeneous nature of the multilamellar vesicles, they provide a binding assay that was rather independent of the curvature. Complementary to the flotation assay, the SMS approach was employed to reveal the curvature preference of proteins.
Finally, we performed the experiment on large GUVs suggested by the referee using CdvB as an example, but this turned out to be inconclusive because the protein forms clusters: these clusters may be creating local curvature at the nanometer scale, which cannot be resolved by optical microscopy (Author response image 4). This is quite typical for proteins that recognize curvature (cf. for instance: De Franceschi N, Alqabandi M, Miguet N, Caillat C, Mangenot S, Weissenhorn W, Bassereau P. The ESCRT protein CHMP2B acts as a diffusion barrier on reconstituted membrane necks. J Cell Sci. 2018 Aug 3;132(4):jcs217968.)
Author response image 4.
Fluorescently labelled CdvB bound to giant unilamellar vesicle. The protein was added in the outer buffer. CdvB forms distinct clusters, which may generate a local region of high membrane curvature.
(2) Line 138-139: How is His-ZipA binding the membrane? Wouldn't Ni2+-NTA lipids be required? If not, how is the binding achieved?
Indeed, NTA-lipids were present. This is now stated both in the legend and in the methods.
(3) In the encapsulated protein assays, why does the luminal fluorescence intensity of the encapsulated protein sometimes appear similar to the bulk fluorescence signal? Since only a small fraction of the protein assembles at membrane necks, shouldn't the luminal pool of unbound protein show higher fluorescence intensity inside the liposomes?
We thank the referee for raising this point and giving us the opportunity to explain this. The reason is that Cdv proteins have a very high affinity for the neck, and when they cluster at the neck the fluorescence intensity of the cluster is many times higher than the background fluorescence. Because we were interested in imaging the clusters and avoiding overexposing them, we adjusted the imaging conditions accordingly, with the result that the fluorescence from both the lumen and the bulk is at very low level.
By choosing different imaging conditions, however, it can be actually seen that the signal inside the lumen is clearly higher than the bulk: this can be seen for instance in Author response image 2, where the brightness has been properly adjusted.
(4) Line 184-185: In Fig. 2I, some CdvB2ΔC puncta seem independent of CdvB1 and are not localized at membrane necks. How many such puncta exist? For example, in the provided micrograph, 2 out of 5 clusters are independent of CdvB1. This proportion is significant. Could the authors quantify the prevalence of these structures and discuss why they form?
We thank the referee for giving us the opportunity to explain this apparent discrepancy. We’ll like to stress the fact that CdvB2ΔC and CdvB1 form an obligate heterodimer: in all our experiments, without exception, we find that they form a strong complex when we mix the two proteins. This is true both in dumbbells and in flotation assays.
In the particular example of Figure 2I, it indeed appears that there are some clusters of CdvB2ΔC that do not contain CdvB1 (we indicated them in Author response image 3 by red arrowheads), while the yellow arrowheads indicate clusters that contain both proteins. It can be clearly seen that the clusters that do contain both proteins (yellow arrows) are localized at necks, while those that only contain CdvB2ΔC (red arrows) are not localized at necks. This is no coincidence. The clusters indicated by the red arrow do contain CdvB1. However, these clusters rapidly diffuse on the membrane plane because they are not fixed at the neck: therefore, they constantly shift in and out of focus. Because there is a time delay in the acquisition of each channel (between 0.5 and 1 second), these cluster were in focus when the CdvB2ΔC signal was being acquired, but sifted out of focus when the CdvB1 signal was being acquired. This implies that the clusters indicated by the yellow arrowheads are stably localized at necks, which is precisely the point we wished to make with this experiment: because Cdv proteins have affinity for curved geometry, they preferentially and stably localize at necks. Why don’t all the clusters localize at necks then?
(5) Figure 1E and 1F: Why do lipids accumulate and colocalize with the proteins? How can the authors confirm lumen connectivity between vesicles? Performing FRAP assays could validate protein localization and enrichment at the lumen of the membrane necks.
At first sight, indeed some lipid enrichment seems to be observed at the neck between lobes of dumbbells.
This is, however, an imaging artifact due to the fact that the neck is diffraction limited. As shown in the Author response image 5, we are acquiring the membrane signal from both lobes at the neck region, and therefore the signal is roughly double, hence the apparent lipid enrichment.
Author response image 5.
Schematic illustrating that the neck between two lobes is smaller than the diffraction limit of optical microscopy (the size of a typical pixel is indicated by the green square). Because of this technical limitation, the fluorescence intensity of the membrane at the neck is twice that of a single membrane.
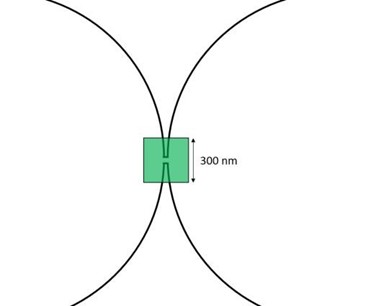
The referee is correct in pointing out that these images do not prove that the lobes are connected, and that FRAP assays is the only way to prove this point. However, in previous papers we have confirmed extensively that in chains of dumbbells the lobes are connected:
- De Franceschi N, Pezeshkian W, Fragasso A, Bruininks BMH, Tsai S, Marrink SJ, Dekker C. Synthetic Membrane Shaper for Controlled Liposome Deformation. ACS Nano. 2022 Nov 28;17(2):966–78. doi: 10.1021/acsnano.2c06125.
- De Franceschi N, Barth R, Meindlhumer S, Fragasso A, Dekker C. Dynamin A as a one-component division machinery for synthetic cells. Nat Nanotechnol. 2024 Jan;19(1):70-76. doi: 10.1038/s41565023-01510-3.
Random sticking of liposomes would also generate clusters of vesicles, not linear chains. We now provide also a Movie (Movie 1) supporting this point.
Investigating whether the necks are open or closed after Cdv reconstitution is indeed a very relevant question, that could be rephrased as “verify whether Cdv proteins or their combination can induce membrane scission”. This is however beyond the scope of this manuscript, as the current work merely addressed the question of hierarchical recruitment of Cdv proteins at the membrane. We plan to examine this in future work.
(6) Why didn't the authors use the same lipid composition, particularly the same proportion of negatively charged lipids, on the SUVs of the flotation assays and on the dumbbell-shaped liposomes?
In flotation assays, it is typical to use a relatively large proportion of negatively charged lipids, to promote protein binding. This is because the aim is to maximize membrane coverage by the protein. The SMS procedure to generate dumbbell-shaped GUVs is completely different, however. Rather than covering the membrane with protein, the idea is to reduce the amount of protein to a minimum, so that any curvature preference can be best visualized. This is e.g. routinely done in tube pulling experiments, for the same reason (See for instance Prévost C, Zhao H, Manzi J, Lemichez E, Lappalainen P, Callan-Jones A, Bassereau P. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 2015 Oct 15;6:8529. doi: 10.1038/ncomms9529).
(7) Line 117-119: The suggestion that polymer formation between CdvA and CdvB facilitates membrane recruitment is intriguing. However, fluorescence microscopy experiments could better elucidate whether there is sequential recruitment of CdvB followed by CdvA, or if these proteins form a heteropolymer composite for membrane binding. Can CdvB bind membranes independently, or does this require synergy between CdvA and CdvB.
We thank the referee for prompting us to perform this experiment. As we now show in Figure 1C, CdvB indeed is able to bind the membrane independently of CdvA. Whether this happens sequentially or simultaneously is an interesting question, but one that is impossible to address with either the SMS or the flotation assay, because in both cases we can only observe the endpoint of the recruitment.
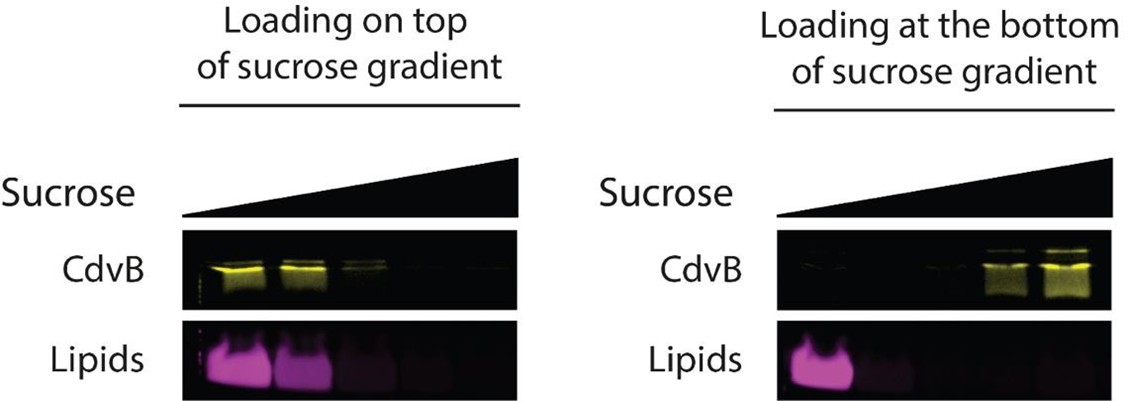
We would also like to clarify one specific experimental detail. Perhaps unsurprisingly, the results from the flotation assay are dependent on the way the assay is performed. In particular, we observed that the same protein can exhibit a different binding profile depending on whether it is being loaded either at the top or at the bottom of the gradient. This can be seen in Author response image 6. This is counterintuitive, since once the equilibrium is reached, the result should only depend on the density of the sample. We performed an overnight centrifugation (> 16 hours) on a short tube (< 3 cm tall), thus equilibrium is being reached (which is corroborated by the fact that CdvB1 and CdvB2 can float to the top of the gradient within this timespan, as shown in Figure 2C, 2E, 2G). We ascribe the difference between top and bottom loading to the fact that, when the sample is loaded at the bottom, it has to be mixed with a concentrated sucrose solution, while in the case of loading from the top, this is not done.
In literature, both loading from top and from bottom have been used:
- Lata S, Schoehn G, Jain A, Pires R, Piehler J, Gottlinger HG, Weissenhorn W. Helical structures of ESCRTIII are disassembled by VPS4. Science. 2008 Sep 5;321(5894):1354-7. doi: 10.1126/science.1161070
- Moriscot C, Gribaldo S, Jault JM, Krupovic M, Arnaud J, Jamin M, Schoehn G, Forterre P, Weissenhorn W, Renesto P. Crenarchaeal CdvA forms double-helical filaments containing DNA and interacts with ESCRT-III-like CdvB. PLoS One. 2011;6(7):e21921. doi: 10.1371/journal.pone.0021921.
- Senju Y, Lappalainen P, Zhao H. Liposome Co-sedimentation and Co-flotation Assays to Study LipidProtein Interactions. Methods Mol Biol. 2021;2251:195-204. doi: 10.1007/978-1-0716-1142-5_14. In performing the flotation assay for CdvB1 and CdvB2ΔC, or when using all 4 proteins together, we loaded the sample at the bottom, and we could detect reproducible binding to liposomes (Figures 2D, 2F, 2H, 3A). However, CdvB does not bind the membrane when loaded at the bottom. Thus, for the experiments shown in figure 1C, we loaded the proteins at the top. This experimental setup allowed us to highlight that CdvB indeed induce a stronger interaction between CdvA and the membrane.
Author response image 6.
CdvB binding to multilamellar vesicles in a flotation assay. In the left panel, the sample was loaded at the top of the sucrose gradient; in the right panel it was loaded at the bottom.
(8) Line 165-173: The authors claim that filament curvature differs between CdvB2ΔC alone and the CdvB1:CdvB2ΔC complex. Are these differences statistically significant? What is the sample size (N)? Furthermore, how do the authors confirm interactions between these proteins in the absence of membranes based solely on EM micrographs?
We can confirm that the filaments are composed by both proteins, because the filaments have different curvature when both proteins are present. However, as requested by referee 3, point (7), we removed the quantification of curvature from panel 2C. We report the N number in the text.
(9) Line 121-123: Are the authors referring to positive or negative membrane curvatures? The cited literature suggests ESCRT-III proteins either lack curvature preferences (e.g., Snf7, CHMP4B) or prefer high positive curvature (e.g., late ESCRT-III subunits). This is confusing since the authors later test recruitment to negatively curved necks.
We do not claim that Cdv proteins prefer positive or negative curvature, because the necks present in dumbbells have a catenoid geometry, which include both positive and negative curvature. We have now clarified this in the discussion.
(10) Since the conclusions rely on the oligomeric state of the proteins, providing SEC-MALS spectra to show the protein oligomeric state right after the purification would strengthen the claims.
While such SEC-MALDI experiments may be interesting, practical implementation of this is not possible since both first authors left the lab on to new positions.
(11) Line 157-160: Suppl. Fig. 2 shows only a single EM micrograph of a small filament. Could the authors provide lower magnification images showing more filaments?
As requested by Referee 3, point (7), we have toned down the importance of these short filaments.
Also, why are the sample sizes for filament length (N=161) and width (N=129) different?
Protein filaments formed by Cdv tend to stick to each other side by side, so that for some filaments the width could not be accurately assessed, and accordingly those were removed from the analysis.
(12) The introduction states that CdvA binds membranes while CdvB does not. However, the results suggest CdvB facilitates membrane binding, helping CdvA attach. This discrepancy needs further explanation.
We thank the referee for raising this point. We have now performed additional experiments (both SMS assay and flotation assays) showing that indeed CdvB from M. sedula is (unlike CdvB from Sulfolobus) able to bind the membrane on its own (Figure 1C, 1F).
Reviewer #2 (Recommendations for the authors):
Best practice would be to show single fluorescence channels in grayscale or inverted grayscale, retaining pseudocolouring only for the merged multichannel image.
We decided to retain and standardize the colors, both for gels and for microscopy images, in order to have the same color-code for each protein. We believe this improves readability, and this was also a request from Referee 3. Thus, throughout the manuscript, CdvA is in grayscale, CdvB in yellow, CdvB1 in green, CdvB2ΔC in cyan and the membrane in magenta.
It would be great to include a quantification of liposome curvature vs focal intensity of the various Cdv components - across figures.
Quantification of liposome curvature at the neck can be done (De Franceschi et al., Nature Nanotech. 2024). However, in practice, this requires transferring of the sample post-preparation into a new chamber in order to increase the signal-to-noise ratio of the encapsulated dye, a procedure that drastically reduces the yield of dumbbells. The very sizeable amount of work required to obtain reliable measurements, especially considering all the proteins and protein combinations used in this study, indicates that this represents a project in itself, which goes well beyond the scope of this manuscript.
Reviewer #3 (Recommendations for the authors):
(1) We would encourage the authors to consider including the length of the scale bar next to the scale bar in each image and not in the figure description. This would greatly aid in clarity and interpretation of figures.
We have now written the length of the scale bar in the figures.
(2) In a similar vein, could the authors consider labeling panels throughout the manuscript, writing that sample is being presented? This goes mainly for the negative stain and the dumbbell fluorescence images, as having to continuously consult the figure legend again hinders clarity.
We have now labelled the EM images as requested by the referee.
(3) Lines 254-256: would the statement hold not only for CdvB2∆C, but for all imaged proteins? They all seem to localize to membrane necks, presumably favoring membrane binding to a specific membrane topology.
We agree with the referee, and changed the phrasing accordingly.
(4) CdvB2∆C construct - presumably this was a truncation of helix 5 of the ESCRT-III domain? Figure 1A shows that the ESCRT-III domain spans residues 34-170 and therefore implies that all five ESCRT-III helices (which make up the ESCRT-III domain) are present in the C-terminal truncation. Could the authors clarify?
Indeed, the truncation was done at residue 170.
(5) Results of the liposome flotation assays are presented inconsistently across the three figures (Figs 1C, 2DFH, and 3A). This makes it more difficult than it needs to be to interpret and compare results. Could the authors consider presenting the three gels in a more similar, standardized way across the three figures?
To improve readability, we now standardized the colors, both for gels and for microscopy images, in order to have the same color-code for each protein. Thus, throughout the manuscript, CdvA is in grayscale, CdvB in yellow, CdvB1 in green, CdvB2ΔC in cyan and the membrane in magenta.
(6) From the data presented in Fig 1EF, it cannot be concluded whether CdvB and CdvA colocalize, as only one protein is labelled. Is there a technical reason for this?
We have now repeated the same experiment by having both proteins labelled, confirming that there is co-localization at the neck (Figure 1G).
(7) Fig 2C: is the difference between the two samples significant
As requested by Referee 3, we have removed Figure 2C.
(8) Fig 2I is missing a 'merged' panel.
We have now added the merged panel.
(9) The fluorescence intensity plots in Supp Figs 1C and 3C would be easier to interpret if the lipid and protein signal would be plotted on the same plot (say, with normalized fluorescence intensity)
It is not immediately obvious to us what the signal should be normalized to. What we wished to convey with these plots was that the intensity of proteins spikes at the neck region. In an attempt to improve clarity, we have now aligned the plots vertically, and highlighted the position of the neck.
(10) CdvA should have a capital "A" in Figure 3A, panel 3.
We have now corrected this.
(11) The discussion doesn't comment on the need to truncate CdvB2.
This is explained in the result session.
-
eLife Assessment
This valuable study investigates how the proteins of the Cdv division system in Metallosphaera sedula archaea sequentially interact with curved membranes in vitro, extending our understanding of this reduced ESCRT-like machinery. While the data support key aspects of protein recruitment and membrane remodeling, missing controls and statistical analysis information, unaddressed discrepancies, and limitations in recapitulating native geometry leave the data incomplete to fully support the proposed conclusions. The work will be of interest to evolutionary and synthetic biologists as membrane biophysicists but would benefit from additional experiments and a more cautious interpretation of results.
-
Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbell-shaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two sub-complexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane necks. Similarly, the findings on CdvB1:CdvB2ΔC indicate that truncation of CdvB2 facilitates …Reviewer #1 (Public review):
Summary:
The authors aimed to elucidate the recruitment order and assembly of the Cdv proteins during Sulfolobus acidocaldarius archaeal cell division using a bottom-up reconstitution approach. They employed liposome-binding assays, EM, and fluorescence microscopy with in vitro reconstitution in dumbbell-shaped liposomes to explore how CdvA, CdvB, and the homologues of ESCRT-III proteins (CdvB, CdvB1, and CdvB2) interact to form membrane remodeling complexes.
The study sought to reconstitute the Cdv machinery by first analyzing their assembly as two sub-complexes: CdvA:CdvB and CdvB1:CdvB2ΔC. The authors report that CdvA binds lipid membranes only in the presence of CdvB and localizes preferentially to membrane necks. Similarly, the findings on CdvB1:CdvB2ΔC indicate that truncation of CdvB2 facilitates filament formation and enhances curvature sensitivity in interaction with CdvB1. Finally, while the authors reconstitute a quaternary CdvA:CdvB:CdvB1:CdvB2 complex and demonstrate its enrichment at membrane necks, the mechanistic details of how these complexes drive membrane remodeling by subcomplexes removal by the proteasome and/or CdvC remain speculative.
Although the work highlights intriguing similarities with eukaryotic ESCRT-III systems and explores unique archaeal adaptations, the conclusions drawn would benefit from stronger experimental validation and a more comprehensive mechanistic framework.Strengths:
The study of machinery assembly and its involvement in membrane remodeling, particularly using bottom-up reconstituted in vitro systems, presents significant challenges. This is particularly true for systems like the ESCRT-III complex, which localizes uniquely at the lumen of membrane necks prior to scission. The use of dumbbell-shaped liposomes in this study provides a promising experimental model to investigate ESCRT-III and ESCRT-III-like protein activity at membrane necks.
The authors present intriguing evidence regarding the sequential recruitment of ESCRT-III proteins in crenarchaea-a close relative of eukaryotes. This finding suggests that the hierarchical recruitment characteristic of eukaryotic systems may predate eukaryogenesis, which is a significant and exciting contribution. However, the broader implications of these findings for membrane remodeling mechanisms remain speculative, and the study would benefit from stronger experimental validation and expanded contextualization within the field.Weaknesses:
This manuscript presents several methodological inconsistencies and lacks key controls to validate its claims. Additionally, there is insufficient information about the number of experimental repetitions, statistical analyses, and a broader discussion of the major findings in the context of open questions in the field.
-
Reviewer #2 (Public review):
Summary:
The Crenarchaeal Cdv division system represents a reduced form of the universal and ubiquitous ESCRT membrane reverse-topology scission machinery, and therefore a prime candidate for synthetic and reconstitution studies. The work here represents a solid extension of previous work in the field, clarifying the order of recruitment of Cdv proteins to curved membranes.
Strengths:
The use of a recently developed approach to produce dumbbell-shaped liposomes (De Franceschi et al. 2022), which allowed the authors to assess recruitment of various Cdv assemblies to curved membranes or membrane necks; reconstitution of a quaternary Cdv complex at a membrane neck.
Weaknesses:
The manuscript is a bit light on quantitative detail, across the various figures, and several key controls are missing (CdvA, B alone to …
Reviewer #2 (Public review):
Summary:
The Crenarchaeal Cdv division system represents a reduced form of the universal and ubiquitous ESCRT membrane reverse-topology scission machinery, and therefore a prime candidate for synthetic and reconstitution studies. The work here represents a solid extension of previous work in the field, clarifying the order of recruitment of Cdv proteins to curved membranes.
Strengths:
The use of a recently developed approach to produce dumbbell-shaped liposomes (De Franceschi et al. 2022), which allowed the authors to assess recruitment of various Cdv assemblies to curved membranes or membrane necks; reconstitution of a quaternary Cdv complex at a membrane neck.
Weaknesses:
The manuscript is a bit light on quantitative detail, across the various figures, and several key controls are missing (CdvA, B alone to better interpret the co-polymerisation phenotypes and establish the true order of recruitment, for example) - addressing this would make the paper much stronger. The authors could also include in the discussion a short paragraph on implications for our understanding of ESCRT function in other contexts and/or in archaeal evolution, as well as a brief exploration of the possible reasons for the discrepancy between the foci observed in their liposome assays and the large rings observed in cells - to better serve the interests of a broad audience.
-
Reviewer #3 (Public review):
Summary:
In this report, De Franceschi et al. purify components of the Cdv machinery in archaeon M. sedula and probe their interactions with membrane and with one-another in vitro using two main assays - liposome flotation and fluorescent imaging of encapsulated proteins. This has the potential to add to the field by showing how the order of protein recruitment seen in cells is related to the differential capacity of individual proteins to bind membranes when alone or when combined.
Strengths:
Using the floatation assay, they demonstrate that CdvA and CdvB bind liposomes when combined. While CdvB1 also binds liposomes under these conditions, in the floatation assay, CdvB2 lacking its C-terminus is not efficiently recruited to membranes unless CdvAB or CdvB1 are present. The authors then employ a clever …
Reviewer #3 (Public review):
Summary:
In this report, De Franceschi et al. purify components of the Cdv machinery in archaeon M. sedula and probe their interactions with membrane and with one-another in vitro using two main assays - liposome flotation and fluorescent imaging of encapsulated proteins. This has the potential to add to the field by showing how the order of protein recruitment seen in cells is related to the differential capacity of individual proteins to bind membranes when alone or when combined.
Strengths:
Using the floatation assay, they demonstrate that CdvA and CdvB bind liposomes when combined. While CdvB1 also binds liposomes under these conditions, in the floatation assay, CdvB2 lacking its C-terminus is not efficiently recruited to membranes unless CdvAB or CdvB1 are present. The authors then employ a clever liposome assay that generates chained spherical liposomes connected by thin membrane necks, which allows them to accurately control the buffer composition inside and outside of the liposome. With this, they show that all four proteins accumulate in necks of dumbbell-shaped liposomes that mimic the shape of constricting necks in cell division. Taken altogether, these data lead them to propose that Cdv proteins are sequentially recruited to the membrane as has also been suggested by in vivo studies of ESCRT-III dependent cell division in crenarchaea.
Weaknesses:
These experiments provide a good starting point for the in vitro study the interaction of Cdv system components with the membrane and their consecutive recruitment. However, several experimental controls are missing that complicate their ability to draw strong conclusions. Moreover, some results are inconsistent across the two main assays which make the findings difficult to interpret.
(1) Missing controls.
Various protein mixtures are assessed for their membrane-binding properties in different ways. However, it is difficult to interpret the effect of any specific protein combination, when the same experiment is not presented in a way that includes separate tests for all individual components. In this sense, the paper lacks important controls.
For example, Fig 1C is missing the CdvB-only control. The authors remark that CdvB did not polymerise (data not shown) but do not comment on whether it binds membrane in their assays. In the introduction, Samson et al., 2011 is cited as a reference to show that CdvB does not bind membrane. However, here the authors are working with protein from a different organism in a different buffer, using a different membrane composition and a different assay. Given that so many variables are changing, it would be good to present how M. sedula CdvB behaves under these conditions.
Similarly, there is no data showing how CdvB alone or CdvA alone behave in the dumbbell liposome assay. Without these controls, it's impossible to say whether CdvA recruits CdvB or the other way around.
The manuscript would be much stronger if such data could be added.
(2) Some of the discrepancies in the data generated using different assays are not discussed.
The authors show that CdvB2∆C binds membrane and localizes to membrane necks in the dumbbell liposome assay, but no membrane binding is detected in the flotation assay. The discrepancy between these results further highlights the need for CdvB-only and CdvA-only controls.
(3) Validation of the liposome assay.
The experimental setup to create dumbbell-shaped liposomes seems great and is a clever novel approach pioneered by the team. Not only can the authors manipulate liposome shape, they also state that this allows them to accurately control the species present on the inside and outside of the liposome. Interpreting the results of the liposome assay, however, depends on the geometry being correct. To make this clearer, it would seem important to include controls to prove that all the protein imaged at membrane necks lie on the inside of liposomes. In the images in SFig3 there appears to be protein outside of the liposome. It would also be helpful to present data to show test whether the necks are open, as suggested in the paper, by using FRAP or some other related technique.
(4) Quantification of results from the liposome assay.
The paper would be strengthened by the inclusion of more quantitative data relating to the liposome assay. Firstly, only a single field of view is shown for each condition. Because of this, the reader cannot know whether this is a representative image, or an outlier? Can the authors do some quantification of the data to demonstrate this? The line scan profiles in the supplemental figures would be an example of this, but again in these Figures only a single image is analyzed.
We would recommend that the authors present quantitative data to show the extent of co-localization at the necks in each case. They also need a metric to report instances in which protein is not seen at the neck, e.g. CdvB2 but not CdvB1 in Fig2I, which rules out a simple curvature preference for CdvB2 as stated in line 182.
Secondly, the authors state that they see CdvB2∆C recruited to the membrane by CdvB1 (lines 184-187, Fig 2I). However, this simple conclusion is not borne out in the data. Inspecting the CdvB2∆C panels of Fig 2I, Fig3C, and Fig3D, CdvB2∆C signal can be seen at positions which don't colocalize with other proteins. The authors also observe CdvB2∆C localizing to membrane necks by itself (Fig 2E). Therefore, while CdvB1 and CdvB2∆C colocalize in the flotation assay, there is no strong evidence for CdvB2∆C recruitment by CdvB1 in dumbbells. This is further underscored by the observation that in the presented data, all Cdv proteins always appear to localize at dumbbell necks, irrespective of what other components are present inside the liposome. Although one nice control is presented (ZipA), this suggests that more work is required to be sure that the proteins are behaving properly in this assay. For example, if membrane binding surfaces of Cdv proteins are mutated, does this lead to the accumulation of proteins in the bulk of the liposome as expected?
(5) Rings.
The authors should comment on why they never observe large Cdv rings in their experiments. In crenarchaeal cell division, CdvA and CdvB have been observed to form large rings in the middle of the 1 micron cell, before constriction. Only in the later stages of division are the ESCRTs localized to the constricting neck, at a time when CdvA is no longer present in the ring. Therefore, if the in vitro assay used by the authors really recapitulated the biology, one would expect to see large CdvAB rings in Figs 1EF. This is ignored in the model. In the proposed model of ring assembly (line 252), CdvAB ring formation is mentioned, but authors do not discuss the fact that they do not observe CdvAB rings - only foci at membrane necks. The discussion section would benefit from the authors commenting on this.
(6) Stoichiometry
It is not clear why 100% of the visible CdvA and 100% of the the visible CdvB are shifted to the lipid fraction in 1C. Perhaps this is a matter of quantification. Can the authors comment on the stoichiometry here?
(7) Significance of quantification of MBP-tagged filaments.
Authors use tagging and removal of MBP as a convenient, controllable system to trigger polymerisation of various Cdv proteins. However, it is unclear what is the value and significance of reporting the width and length of the short linear filaments that are formed by the MBP-tagged proteins. Presumably they are artefactual assemblies generated by the presence of the tag? Similar Figure 2C doesn't seem a useful addition to the paper.
-
Author response:
We thank the three Reviewers for the extensive evaluation of our work, which was largely positive and constructive. Prompted by their reviews and the many suggestions, we plan to do additional control experiments to add further data in a revised manuscript in order to improve the statistics and quantitation. Furthermore, we plan to expand the discussion. We agree that a more comprehensive mechanistic framework would be welcome but note that the system is a complex multicomponent system which is challenging. We plan to expand the work in future follow-up research.
-
-
