Spatial tuning of face part representations within face-selective areas revealed by high-field fMRI
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
How the brain is organized to represent various concepts has long been a central cognitive neuroscience research topic. Zhang and colleagues investigated the spatial distribution of feature tuning for different face-parts within face-selective regions of human visual cortex using ultra-high resolution 7.0 T fMRI. The findings complement non-human primate studies of face-selective patches and will be of interest to psychologists and system neuroscientists.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #3 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Regions sensitive to specific object categories as well as organized spatial patterns sensitive to different features have been found across the whole ventral temporal cortex (VTC). However, it is unclear that within each object category region, how specific feature representations are organized to support object identification. Would object features, such as object parts, be represented in fine-scale spatial tuning within object category-specific regions? Here, we used high-field 7T fMRI to examine the spatial tuning to different face parts within each face-selective region. Our results show consistent spatial tuning of face parts across individuals that within right posterior fusiform face area (pFFA) and right occipital face area (OFA), the posterior portion of each region was biased to eyes, while the anterior portion was biased to mouth and chin stimuli. Our results demonstrate that within the occipital and fusiform face processing regions, there exist systematic spatial tuning to different face parts that support further computation combining them.
Article activity feed
-

-

Author Response:
Reviewer #2:
In Zhang et al.'s paper, with 7T fMRI, they used different face parts as stimuli to explore the functional organization within the face specific areas, and found consistent patterns between different subjects in rFFA and rOFA. In these areas, the posterior region was biased to eye, and the anterior region was biased to mouth. To exclude potential confounds, they also ran several control experiments to show that the preference to eyes and mouth is not due to the eccentricity or upper-lower visual field preference. Based on what they found, they claim that there exists a finer scale functional organization within the face areas.
In general, I think the whole study is carefully designed, and the results are solid and interesting. However, I am not very comfortable about the claim about the organization of …
Author Response:
Reviewer #2:
In Zhang et al.'s paper, with 7T fMRI, they used different face parts as stimuli to explore the functional organization within the face specific areas, and found consistent patterns between different subjects in rFFA and rOFA. In these areas, the posterior region was biased to eye, and the anterior region was biased to mouth. To exclude potential confounds, they also ran several control experiments to show that the preference to eyes and mouth is not due to the eccentricity or upper-lower visual field preference. Based on what they found, they claim that there exists a finer scale functional organization within the face areas.
In general, I think the whole study is carefully designed, and the results are solid and interesting. However, I am not very comfortable about the claim about the organization of the face areas. Typically, when we talk about the organization, it either has more than 2 subdivisions or it has a continuous representation of certain features. In this paper, the results are mainly about the comparison between two face parts, and they failed to find other distinctive subareas showing preference to other face parts. Therefore, I would suggest that the authors could tune down their claim from functional organization to functional preference.
We have followed the advice from the reviewer to tune down the claim of functional organization in our manuscript. To emphasize both the functional preferences to different face parts within face-selective regions and the consistent spatial profile across different individuals, we now use “spatial tuning of face parts” in the manuscript.
Reviewer #3:
Zhang and colleagues investigated the spatial distribution of feature tuning for different face-parts within face-selective regions of human visual cortex using ultra-high resolution 7.0 T fMRI. By comparing the response patterns elicited by images of face-parts (hair, eyes, nose, mouth and chin) with whole faces, they report a spatial pattern of tuning for eyes and mouth along the posterior-anterior axis of both the pFFA and OFA. Within the pFFA this pattern spatial tuning appeared to track the orientation of the mid fusiform sulcus - an anatomical landmark for face-processing in ventral temporal cortex. Two additional control experiments are conducted to examine the robustness of the original findings and to rule out potentially confounding variables. These data are consistent with recent evidence for similar face-part tuning in the OFA and add to the growing body of work showing the topographical mapping feature based tuning within visual cortex.
The conclusions of this paper are mostly supported by the data, but some aspects of the data acquisition, analysis and interpretation that require further clarification/consideration.
- It is currently unclear whether the current data are in full agreement with recent work (de Haas et al., 2021) showing similar face-part tuning within the OFA (or IOG) bilaterally. The current data suggest that feature tuning for eye and mouth parts progresses along the posterior-anterior axis within the right pFFA and right OFA. In this regard, the data are consistent. But de Haas and colleagues also demonstrated tuning for visual space that was spatially correlated (i.e. upper visual field representations overlapped upper face-part preferences and vice-versa). The current manuscript found little evidence for this correspondence within pFFA but does not report the data for OFA. For completeness this should be reported and any discrepancies with either the prior, or between OFA and pFFA discussed.
In the current study, three participants had data from both retinotopic mapping and face part mapping experiments. Consistent and robust part clustering were found in the right pFFA and right OFA. Following the reviewer’s suggestion, we analyzed these data for the right OFA and found the spatial patterns of eyes vs. mouths are similar to the patterns of visual field sensitivity on the vertical direction (i.e., upper to lower visual field), which are consistent with de Haas and colleagues’ findings. Note that we used more precise functional localization of OFA, while de Haas et al’s analysis was based on anatomically defined IOG, for which OFA is a part of. We have added this result in the Results session (Page 16), and also added a supplemental Figure 4-figure supplement 1.
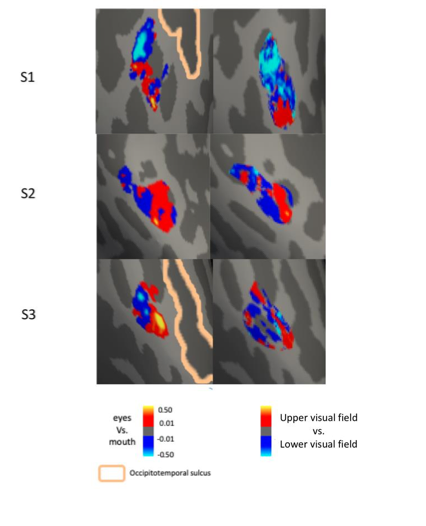
- It is somewhat challenging to fully interpret the responses to face-parts when they were presented at fixation and not in the typical visual field locations during real-world perception. For instance, we typically fixate faces either on or just below the eyes (Peterson et al., 2012) and so in the current experiment the eyes are in the typical viewing position, but the remainder of the face-parts are not (e.g. when fixating the eyes, the nose mouth and chin all fall in the lower visual field but in the current experimental paradigm they appear at fixation). Consideration of whether the reported face-part tuning would hold (or even be enhanced) if face-parts were presented in their typical locations should be included.
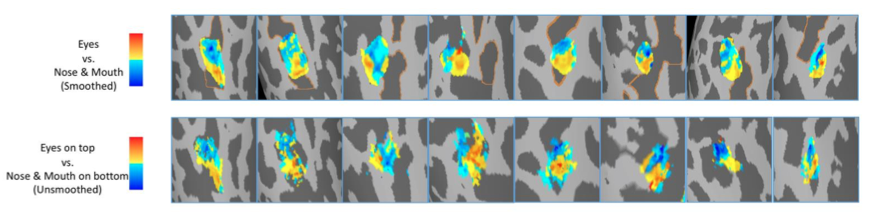
Our early visual cortex and some of the object-selective visual areas are sensitive to visual field locations. To dissociate the visual field tuning and face part tuning in face processing regions, in the main experiment of the current study the face part stimuli were presented at fixation to avoid the potential confounding contribution from visual field location. The spatial correlation between face part tuning and visual field tuning has been observed in posterior part of the face network. It is unlikely that presenting the face parts at the fixation was responsible for the observed face part tuning. To directly test the role of stimulus location, we reanalyzed the data from control experiment 2 in which face parts were presented at their typical locations. Contrasting eyes above fixation vs. nose & mouth below fixation revealed similar anterior-posterior bias in the right pFFA, showing that the face part tuning in the right pFFA is invariant to the visual field location of stimuli. See comparison in the figure below, note that the maps of eyes on top vs. nose & mouth on bottom are unsmoothed:
- Although several experiments (including two controls) have been conducted, each one runs the risk of being underpowered (n ranges 3-10). One way to add reassurance when sample sizes are small is to include analyses of the reliability and replicability of the data within subjects through a split-half, or other cross-validation procedure. The main experiment here consisted of eight functional runs, which is more than sufficient for these types of analyses to be performed.
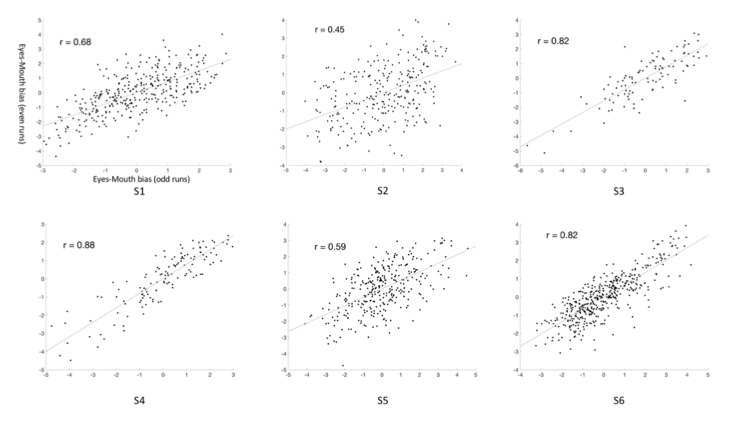
Following the reviewer’s suggestion, we split the eight runs data from each participant in the main experiment into two data sets (odd-runs and even-runs), and estimated the eyes-mouth biases within each data set. Then we calculated the correlation coefficient between such biases across different voxels between the two data sets to estimate the reliability of the results in the right pFFA. The results demonstrate strong reliability of the data within participants. We have added these results in the Results session (Page 7 and Figure 2-figure supplement 1).
- The current findings were only present within the right pFFA and right OFA. Although right lateralisation of face-processing is mentioned in the discussion, this is only cursory. A more expansive discussion of what such a face-part tuning might mean for our understanding of face-processing is warranted, particularly given that the recent work by de Haas and colleagues was bilateral.
The right lateralization of face-processing has been observed in face-selective network. Both the neural selectivity to faces (Kanwisher et al., 1997) and the decodable neural information of faces (Zhang et al., 2015) are higher in the right than in the left hemisphere. The neural clustering of face part tuning and consistent spatial patterns across individuals in the right rather than in the left face selective regions provides a potential computational advantage for right lateralization for face processing. The clustering of neurons with similar feature tuning have been found extensively in the ventral pathway, which may help to support a more efficient neural processing. Therefore, one of the neural mechanisms underlying the functional lateralization of face processing could be the existence of spatial clustering of face part tunings in the right hemisphere. We have added more discussion about the relevance between our results and lateralization of face processing.
-
Evaluation Summary:
How the brain is organized to represent various concepts has long been a central cognitive neuroscience research topic. Zhang and colleagues investigated the spatial distribution of feature tuning for different face-parts within face-selective regions of human visual cortex using ultra-high resolution 7.0 T fMRI. The findings complement non-human primate studies of face-selective patches and will be of interest to psychologists and system neuroscientists.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #3 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
How the brain is organized to represent various concepts has long been a central cognitive neuroscience research topic. Most notably, several areas in occipito-temporal cortex are known to be specialized in representing faces. However, as a type of complex stimuli, faces contain multi-dimension features. By using high-field 7T fMRI, the present paper aimed to examine how face-selective brain areas might be organized to represent face features/face parts in fine-scaled spatial patterns.
This is an overall well-executed study with a careful experimental design that includes helpful control conditions, although the external validity might be somewhat limited due to the small sample size of the present study. Nonetheless, the within-subject results appear to be reliable and the results support their conclusions. …
Reviewer #1 (Public Review):
How the brain is organized to represent various concepts has long been a central cognitive neuroscience research topic. Most notably, several areas in occipito-temporal cortex are known to be specialized in representing faces. However, as a type of complex stimuli, faces contain multi-dimension features. By using high-field 7T fMRI, the present paper aimed to examine how face-selective brain areas might be organized to represent face features/face parts in fine-scaled spatial patterns.
This is an overall well-executed study with a careful experimental design that includes helpful control conditions, although the external validity might be somewhat limited due to the small sample size of the present study. Nonetheless, the within-subject results appear to be reliable and the results support their conclusions. The findings complement non-human primate studies on face-selective patches. In addition, there are some agreements but also discrepancies between the present findings and a previously proposed "faciotopy" hypothesis, i.e., the spatial organization of patches selective to different face parts would reflect the actual distances between these face parts.
-
Reviewer #2 (Public Review):
In Zhang et al.'s paper, with 7T fMRI, they used different face parts as stimuli to explore the functional organization within the face specific areas, and found consistent patterns between different subjects in rFFA and rOFA. In these areas, the posterior region was biased to eye, and the anterior region was biased to mouth. To exclude potential confounds, they also ran several control experiments to show that the preference to eyes and mouth is not due to the eccentricity or upper-lower visual field preference. Based on what they found, they claim that there exists a finer scale functional organization within the face areas.In general, I think the whole study is carefully designed, and the results are solid and interesting. However, I am not very comfortable about the claim about the organization of …
Reviewer #2 (Public Review):
In Zhang et al.'s paper, with 7T fMRI, they used different face parts as stimuli to explore the functional organization within the face specific areas, and found consistent patterns between different subjects in rFFA and rOFA. In these areas, the posterior region was biased to eye, and the anterior region was biased to mouth. To exclude potential confounds, they also ran several control experiments to show that the preference to eyes and mouth is not due to the eccentricity or upper-lower visual field preference. Based on what they found, they claim that there exists a finer scale functional organization within the face areas.In general, I think the whole study is carefully designed, and the results are solid and interesting. However, I am not very comfortable about the claim about the organization of the face areas. Typically, when we talk about the organization, it either has more than 2 subdivisions or it has a continuous representation of certain features. In this paper, the results are mainly about the comparison between two face parts, and they failed to find other distinctive subareas showing preference to other face parts. Therefore, I would suggest that the authors could tune down their claim from functional organization to functional preference.
-
Reviewer #3 (Public Review):
Zhang and colleagues investigated the spatial distribution of feature tuning for different face-parts within face-selective regions of human visual cortex using ultra-high resolution 7.0 T fMRI. By comparing the response patterns elicited by images of face-parts (hair, eyes, nose, mouth and chin) with whole faces, they report a spatial pattern of tuning for eyes and mouth along the posterior-anterior axis of both the pFFA and OFA. Within the pFFA this pattern spatial tuning appeared to track the orientation of the mid fusiform sulcus - an anatomical landmark for face-processing in ventral temporal cortex. Two additional control experiments are conducted to examine the robustness of the original findings and to rule out potentially confounding variables. These data are consistent with recent evidence for …
Reviewer #3 (Public Review):
Zhang and colleagues investigated the spatial distribution of feature tuning for different face-parts within face-selective regions of human visual cortex using ultra-high resolution 7.0 T fMRI. By comparing the response patterns elicited by images of face-parts (hair, eyes, nose, mouth and chin) with whole faces, they report a spatial pattern of tuning for eyes and mouth along the posterior-anterior axis of both the pFFA and OFA. Within the pFFA this pattern spatial tuning appeared to track the orientation of the mid fusiform sulcus - an anatomical landmark for face-processing in ventral temporal cortex. Two additional control experiments are conducted to examine the robustness of the original findings and to rule out potentially confounding variables. These data are consistent with recent evidence for similar face-part tuning in the OFA and add to the growing body of work showing the topographical mapping feature based tuning within visual cortex.
The conclusions of this paper are mostly supported by the data, but some aspects of the data acquisition, analysis and interpretation that require further clarification/consideration.
It is currently unclear whether the current data are in full agreement with recent work (de Haas et al., 2021) showing similar face-part tuning within the OFA (or IOG) bilaterally. The current data suggest that feature tuning for eye and mouth parts progresses along the posterior-anterior axis within the right pFFA and right OFA. In this regard, the data are consistent. But de Haas and colleagues also demonstrated tuning for visual space that was spatially correlated (i.e. upper visual field representations overlapped upper face-part preferences and vice-versa). The current manuscript found little evidence for this correspondence within pFFA but does not report the data for OFA. For completeness this should be reported and any discrepancies with either the prior, or between OFA and pFFA discussed.
It is somewhat challenging to fully interpret the responses to face-parts when they were presented at fixation and not in the typical visual field locations during real-world perception. For instance, we typically fixate faces either on or just below the eyes (Peterson et al., 2012) and so in the current experiment the eyes are in the typical viewing position, but the remainder of the face-parts are not (e.g. when fixating the eyes, the nose mouth and chin all fall in the lower visual field but in the current experimental paradigm they appear at fixation). Consideration of whether the reported face-part tuning would hold (or even be enhanced) if face-parts were presented in their typical locations should be included.
Although several experiments (including two controls) have been conducted, each one runs the risk of being underpowered (n ranges 3-10). One way to add reassurance when sample sizes are small is to include analyses of the reliability and replicability of the data within subjects through a split-half, or other cross-validation procedure. The main experiment here consisted of eight functional runs, which is more than sufficient for these types of analyses to be performed.
The current findings were only present within the right pFFA and right OFA. Although right lateralisation of face-processing is mentioned in the discussion, this is only cursory. A more expansive discussion of what such a face-part tuning might mean for our understanding of face-processing is warranted, particularly given that the recent work by de Haas and colleagues was bilateral.
-
