Neural dynamics of visual working memory representation during sensory distraction
Curation statements for this article:-
Curated by eLife

eLife Assessment
This important study reports a reanalysis of one experiment of a previously-published report to characterize the dynamics of neural population codes during visual working memory in the presence of distracting information. This paper presents solid evidence that working memory representations are dynamic and distinct from sensory representations of intervening distractions. This research will be of interest to cognitive neuroscientists working on the neural bases of visual perception and memory.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Recent studies have provided evidence for the concurrent encoding of sensory percepts and visual working memory (VWM) contents across visual areas; however, it has remained unclear how these two types of representations are concurrently present. Here, we reanalyzed an open-access fMRI dataset where participants memorized a sensory stimulus while simultaneously being presented with sensory distractors. First, we found that the VWM code in several visual regions did not fully generalize between different time points, suggesting a dynamic code. A more detailed analysis revealed that this was due to shifts in coding spaces across time. Second, we collapsed neural signals across time to assess the degree of interference between VWM contents and sensory distractors, specifically by testing the alignment of their encoding spaces. We find that VWM and feature-matching sensory distractors are encoded in coding spaces that do not fully overlap, but the separation decreases when distractors negatively impact behavioral performance in recalling the target. Together, these results indicate a role of dynamic coding and temporally stable coding spaces in helping multiplex perception and VWM within visual areas.
Article activity feed
-

-
-

eLife Assessment
This important study reports a reanalysis of one experiment of a previously-published report to characterize the dynamics of neural population codes during visual working memory in the presence of distracting information. This paper presents solid evidence that working memory representations are dynamic and distinct from sensory representations of intervening distractions. This research will be of interest to cognitive neuroscientists working on the neural bases of visual perception and memory.
-
Reviewer #1 (Public review):
Summary:
In this study, the authors re-analyzed a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly-oriented grating). In this manuscript, the authors focused on identifying whether the neural code in retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but encoded in an …
Reviewer #1 (Public review):
Summary:
In this study, the authors re-analyzed a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly-oriented grating). In this manuscript, the authors focused on identifying whether the neural code in retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but encoded in an alternative format. They identify some timepoints - especially towards the beginning/end of the delay - where the multivariate activation pattern fails to generalize to other timepoints, and interpret this as evidence for a dynamic code. Additionally, the authors compare the representational format of remembered orientation in the presence vs absence of a distracting stimulus, averaged over the delay period. This analysis suggested a 'rotation' of the representational subspace between distracting orientations and remembered orientations, which may help preserve simultaneous representations of both remembered and viewed stimuli. Intriguingly, this rotation was a bit smaller for Expt 2, in which the orientation distractor had a greater behavioral impact on the participants' behavioral working memory recall performance, suggesting that more separation between subspaces is critical for preserving intact working memory representations.
Strengths:
(1) Direct comparisons of coding subspaces/manifolds between timepoints, task conditions, and experiments is an innovative and useful approach for understanding how neural representations are transformed to support cognition
(2) Re-use of existing dataset substantially goes beyond the authors' previous findings by comparing geometry of representational spaces between conditions and timepoints, and by looking explicitly for dynamic neural representations
(3) Simulations testing whether dynamic codes can be explained purely by changes in data SNR are an important contribution, as this rules out a category of explanations for the dynamic coding results observed
Weaknesses:
(1) Primary evidence for 'dynamic coding', especially in early visual cortex, appears to be related to the transition between encoding/maintenance and maintenance/recall, but the delay period representations seem overall stable, consistent with some previous findings. However, given the simulation results, the general result that representations may change in their format appears solid, though the contribution of different trial phases remains important for considering the overall result.
(2) Converting a continuous decoding metric (angular error) to "% decoding accuracy" serves to obfuscate the units of the actual results. Decoding precision (e.g., sd of decoding error histogram) would be more interpretable and better related to both the previous study and behavioral measures of WM performance.
Comments on revised version:
The authors have addressed all my previous concerns.
-
Author response:
The following is the authors’ response to the previous reviews
Reviewer #1:
Reviewer #1 (Recommendations For The Authors):
(1) At several places in the reply to reviewers and the manuscript, when discussing the new simulations conducted, the authors mention they break the 180 trials into a train/test split of 108/108 - is this value correct? If so, how? (pg 19 of updated manuscript)
Thank you for pointing this out; it was not clearly explained. We have now added the explanation to the Methods section:
“For each iteration, we randomly selected 108 responses from the full set of 180 for training, and then independently sampled another 108 from the same full set for testing. This ensured that the same orientation could appear in both sets, consistent with the structure of the original experiment.”
(2) I appreciate the …
Author response:
The following is the authors’ response to the previous reviews
Reviewer #1:
Reviewer #1 (Recommendations For The Authors):
(1) At several places in the reply to reviewers and the manuscript, when discussing the new simulations conducted, the authors mention they break the 180 trials into a train/test split of 108/108 - is this value correct? If so, how? (pg 19 of updated manuscript)
Thank you for pointing this out; it was not clearly explained. We have now added the explanation to the Methods section:
“For each iteration, we randomly selected 108 responses from the full set of 180 for training, and then independently sampled another 108 from the same full set for testing. This ensured that the same orientation could appear in both sets, consistent with the structure of the original experiment.”
(2) I appreciate the authors have added the variance explained of principal components to the axes of Fig. 3, though it took me a while to notice this, and this isn't described in the figure caption at all. It would likely help readers to directly explain what the % means on each axis of Fig. 3.
Thank you, we have now added a description in both Fig. 2 and 3:
“The axes represent the first two principal components, with labels indicating the percent of total explained variance.”
(3) I believe there is a typo/missing word in the new paragraph on pg 15: "neural visual WM representations in the early visual cortices are [[biased]] towards distractors" (I think the bracketed word may be omitted as a typo)
Thank you - fixed.
-
-
-
eLife Assessment
This important study reports a reanalysis of one experiment of a previously-published report to characterize the dynamics of neural population codes during visual working memory in the presence of distracting information. This paper presents solid evidence that working memory representations are dynamic and distinct from sensory representations of intervening distractions. This research will be of interest to cognitive neuroscientists working on the neural bases of visual perception and memory.
-
Reviewer #1 (Public review):
Summary:
In this study, the authors re-analyzed a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly-oriented grating). In this manuscript, the authors focused on identifying whether the neural code in retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but encoded in an …
Reviewer #1 (Public review):
Summary:
In this study, the authors re-analyzed a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly-oriented grating). In this manuscript, the authors focused on identifying whether the neural code in retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but encoded in an alternative format. They identify some timepoints - especially towards the beginning/end of the delay - where the multivariate activation pattern fails to generalize to other timepoints, and interpret this as evidence for a dynamic code. Additionally, the authors compare the representational format of remembered orientation in the presence vs absence of a distracting stimulus, averaged over the delay period. This analysis suggested a 'rotation' of the representational subspace between distracting orientations and remembered orientations, which may help preserve simultaneous representations of both remembered and viewed stimuli. Intriguingly, this rotation was a bit smaller for Expt 2, in which the orientation distractor had a greater behavioral impact on the participants' behavioral working memory recall performance, suggesting that more separation between subspaces is critical for preserving intact working memory representations.
Strengths:
(1) Direct comparisons of coding subspaces/manifolds between timepoints, task conditions, and experiments is an innovative and useful approach for understanding how neural representations are transformed to support cognition
(2) Re-use of existing dataset substantially goes beyond the authors' previous findings by comparing geometry of representational spaces between conditions and timepoints, and by looking explicitly for dynamic neural representations
(3) Simulations testing whether dynamic codes can be explained purely by changes in data SNR are an important contribution, as this rules out a category of explanations for the dynamic coding results observed
Weaknesses:
(1) Primary evidence for 'dynamic coding', especially in early visual cortex, appears to be related to the transition between encoding/maintenance and maintenance/recall, but the delay period representations seem overall stable, consistent with some previous findings. However, given the simulation results, the general result that representations may change in their format appears solid, though the contribution of different trial phases remains important for considering the overall result.
(2) Converting a continuous decoding metric (angular error) to "% decoding accuracy" serves to obfuscate the units of the actual results. Decoding precision (e.g., sd of decoding error histogram) would be more interpretable and better related to both the previous study and behavioral measures of WM performance.
-
Reviewer #2 (Public review):
Summary:
In this work, Degutis and colleagues addressed an interesting issue related to the concurrent coding of sensory percepts and visual working memory contents in visual cortices. They used generalization analyses to test whether working memory representations change over time, diverge from sensory percepts, and vary across distraction conditions. Temporal generalization analysis demonstrated that off-diagonal decoding accuracies were lower than on-diagonal decoding accuracies, regardless of the presence of intervening distractions, implying that working memory representations can change over time. They further showed that the coding space for working memory contents showed subtle but statistically significant changes over time, potentially explaining the impaired off-diagonal decoding performance. The …
Reviewer #2 (Public review):
Summary:
In this work, Degutis and colleagues addressed an interesting issue related to the concurrent coding of sensory percepts and visual working memory contents in visual cortices. They used generalization analyses to test whether working memory representations change over time, diverge from sensory percepts, and vary across distraction conditions. Temporal generalization analysis demonstrated that off-diagonal decoding accuracies were lower than on-diagonal decoding accuracies, regardless of the presence of intervening distractions, implying that working memory representations can change over time. They further showed that the coding space for working memory contents showed subtle but statistically significant changes over time, potentially explaining the impaired off-diagonal decoding performance. The neural coding of sensory distractions instead remained largely stable. Generalization analyses between target and distractor codes showed overlaps but were not identical. Cross-condition decodings had lower accuracies compared to within-condition decodings. Finally, within-condition decoding revealed more reliable working memory representations in the condition with intervening random noises compared to cross-condition decoding using a trained classifier on data from the no-distraction condition, indicating a change in the VWM format between the noise distractor and no-distractor trials.
Strengths:
This paper demonstrates a clever use of generalization analysis to show changes in the neural codes of working memory contents across time and distraction conditions. It provides some insights into the differences between representations of working memory and sensory percepts, and how they can potentially coexist in overlapping brain regions.
Comments on revisions:
I appreciate the authors' efforts in addressing my previous concerns. The inclusion of additional analyses and data has strengthened the paper. I have no further concerns.
-
Author response:
The following is the authors’ response to the original reviews
Reviewer #1:
Weaknesses:
(1) Only Experiment 1 of Rademaker et al (2019) is reanalyzed. The previous study included another experiment (Expt 2) using different types of distractors which did result in distractor-related costs to neural and behavioral measures of working memory. The Rademaker et al (2019) study uses these two results to conclude that neural WM representations are protected from distraction when distraction does not impact behavior, but conditions that do impact behavior also impact neural WM representations. Considering this previous result is critical for relating the present manuscript's results to the previous findings, it seems necessary to address Experiment 2's data in the present work
We thank the reviewer for the proposal to analyze …
Author response:
The following is the authors’ response to the original reviews
Reviewer #1:
Weaknesses:
(1) Only Experiment 1 of Rademaker et al (2019) is reanalyzed. The previous study included another experiment (Expt 2) using different types of distractors which did result in distractor-related costs to neural and behavioral measures of working memory. The Rademaker et al (2019) study uses these two results to conclude that neural WM representations are protected from distraction when distraction does not impact behavior, but conditions that do impact behavior also impact neural WM representations. Considering this previous result is critical for relating the present manuscript's results to the previous findings, it seems necessary to address Experiment 2's data in the present work
We thank the reviewer for the proposal to analyze Experiment 2 where subjects completed the same type of visual working memory task, but instead had either a flashing orientation distractor or a naturalistic (gazebo or face) distractor present during two-thirds of the trials. As the reviewer points out, unlike Experiment 1, these two conditions in Experiment 2 had a behavioral impact on recall accuracy, when compared to the blank delay. We have now run the temporal cross-decoding analysis, temporally-stable neural subspace analysis, and condition cross-decoding analysis in Experiment 2. The results from the stable subspace analysis are present in Figure 3, while the results from the temporal cross-decoding analysis and condition cross-decoding analysis are present in the Supplementary Data.
First, we are unable to draw strong conclusions from the temporal cross-decoding analysis, as the decoding accuracies across time in Experiment 2 are much lower compared to Experiment 1. In some ROIs of the naturalistic distractor condition we see that some diagonal elements are not part of the above-chance decoding cluster, making it difficult to draw any conclusions regarding dynamic clusters. We do see some dynamic coding in the naturalistic condition in V3 where the off-diagonals do not show above-chance decoding. Since the temporal cross-decoding provides low accuracies, we do not examine the dynamics of neural subspaces across time.
We do, however, run the stable subspace analysis on the flashing orientation distractor condition. Just like in Experiment 1, we examine temporally stable target and distractor subspaces. When projecting the distractor onto the working memory target subspace, we see a higher overlap between the two as compared to Experiment 1. A similar pattern is seen also when projecting the target onto the distractor subspace. We still see an above-chance principal angle between the target and distractor; however, this angle is qualitatively smaller compared to Experiment 1. This shows that the degree of separation between the two neural subspaces is impacted by behavioral performance during recall.
(2) Primary evidence for 'dynamic coding', especially in the early visual cortex, appears to be related to the transition between encoding/maintenance and maintenance/recall, but the delay period representations seem overall stable, consistent with previous findings
We agree with the reviewer that we primarily see dynamic coding between the encoding/maintenance and at the end of the maintenance periods, implying the WM representations are stable in most ROIs. The only place where we argue that we might see more dynamic coding during the delay itself is in V1 during the noise distractor trials in Experiment 1.
(3) Dynamicism index used in Figure 1f quantifies the proportion of off-diagonal cells with significant differences in decoding performance from the diagonal cell. It's unclear why the proportion of time points is the best metric, rather than something like a change in decoding accuracy. This is addressed in the subsequent analysis considering coding subspaces, but the utility of the Figure 1f analysis remains weakly justified.
We agree that other metrics can also provide a summary of dynamics; here, the dynamicism index just acts as a summary visualizing the dynamic elements. It offers an intuitive way to visualize peaks and troughs of the dynamic code across the extent of the trial.
(4) There is no report of how much total variance is explained by the two PCs defining the subspaces of interest in each condition, and timepoint. It could be the case that the first two principal components in one condition (e.g., sensory distractor) explain less variance than the first two principal components of another condition.
We thank the reviewer for this comment. We have now included the percent variance explained for the two PCs in both the temporally-stable target and distractor subspace and the dynamic subspace analysis. The percent-explained is comparable across analyses; the first PC ranges from 43-50% and the second ranges from 28-37%. The PCs within each analysis (dynamic no-distractor, orientation and noise distractor; temporally-stable target and distractor) are even closer in range (Figure 2c and 3d).
(5) Converting a continuous decoding metric (angular error) to "% decoding accuracy" serves to obfuscate the units of the actual results. Decoding precision (e.g., sd of decoding error histogram) would be more interpretable and better related to both the previous study and behavioral measures of WM performance.
We thank the reviewer for the comments. FCA is a linear function of the angular error that uses the following equation:
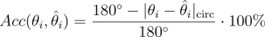
We think that the FCA does not obfuscate the results, but instead provides an intuitive scale where 0% accuracy corresponds to a 180° error, 50% to a 90° error and so on. This also makes it easy to reverse-calculate the absolute error if need be. Our lab has previously used this method in other neuroimaging papers with continuous variables (Barbieri et al. 2023, Weber et al. 2024).
We do, however, agree that “% decoding accuracy” does not provide an accurate reflection of the metric used. We have thus now changed “% decoding accuracy” to “Accuracy (% FCA)”.
(6) This report does not make use of behavioral performance data in the Rademaker et al (2019) dataset.
We have now analyzed Experiment 2 which, as previously mentioned by the reviewer and unlike Experiment 1, showed a decrease in recall accuracy during the two distractor conditions. We address the results from Experiment 2 in a previous response (please see Weaknesses 1).
We do not, however, relate single subject behavioral performance to neural measurements, as we do not think there is enough power to do so with a small number of subjects in both Experiment 1 and 2.
(7) Given there were observed differences between individual retinotopic ROIs in the temporal cross-decoding analyses shown in Figure 1, the lack of data presented for the subspace analyses for the corresponding individual ROIs is a weakness
We have now included an additional supplementary figure that shows individual plots of each ROI for the temporally stable subspace analysis for both Experiment 1 and Experiment 2 (Supplementary Figure 5).
Reviewer #1 (Recommendations For The Authors):
(1) Is there any relationship between stable/dynamic coding properties and aspects of behavioral performance? This seems like a major missed opportunity to better understand the behavioral relevance or importance of the proposed dynamic and orthogonal coding schemes. For example, is it the case that participants who have more orthogonal coding subspaces between orientation distractor and remembered orientation show less of a behavioral consequence to distracting orientations? Less induced bias? I know these differences weren't significant at the group level in the original study, but maybe individual variability in the metrics of this study can explain differences in performance between participants in the reported dataset
As mentioned in the previous response, we do not run individual correlations between dynamic or orthogonal coding metrics and behavioral performance, because of the small number of subjects in both experiments. We believe that for a brain-behavior correlation between average behavioral error of subjects and an average brain measure, we would need a larger sample size.
(2) The voxel selection procedure differs from the original study. The authors should add additional detail about the number of voxels included in their analyses, and how this number of voxels compares to that used in the original study.
We have now added a figure summarizing the number of voxels selected across participants. We do select fewer voxels compared to Rademaker et al. 2019 (see their Supplementary Tables 9 and 10 and our Supplementary Figure 8). For example we have ~500 voxels on average in V1 in Experiment 1, while the original study had ~1000. As mentioned in the methods, we aimed to select voxels that reliably responded to both the perception localizer conditions and the working memory trials.
(3) Lines 428-436 specify details about how data is rescaled prior to decoding. The procedure seems to estimate rescaling factors according to some aspect of the training data, and then apply this rescaling to the training and testing data. Is there a possibility of leakage here? That is - do aspects of the training data impact aspects of the testing data, and could a decoder pick up on such leakage to change decoding? It seems this is performed for each training/testing timepoint pair, and so the temporal unfolding of results may depend on this analysis choice.
Thank you for the suggestion. To prevent data leakage, the mean and standard deviation are computed exclusively from the training set. These scaling parameters are then applied to the test set, ensuring that no information from the test set influences the training process. This transformation simply adjusts the test set to the same scale as the training data, without exposing the model to unseen test data during training.
(4) Figure 1d, V1: it looks like the 'dynamics' are a bit non-symmetric - perhaps the authors could comment on this detail of the results? Why would we expect there would be a dynamic cluster on one side of the diagonal, but not the other? Given that this region, condition is the primary evidence for a dynamic code that's not related to the beginning/end of delay (see other comments), figuring this out is of particular importance.
We thank the reviewer for this question. We think that this is just due to small numerical differences in the upper and lower triangles of the matrix, rather than a neuroscientifically interesting effect. However, this is only a speculative observation.
(5) I think it's important to address the issue I raised in "weaknesses" about variance explained by the top N principal components in each condition. What are we supposed to learn from data projected into subspaces fit to different conditions if the subspaces themselves are differently useful?
Thank you, this has now been addressed in a previous comment (please see Weakness 4).
Reviewer #2:
Weaknesses:
(1) An alternative interpretation of the temporal dynamic pattern is that working memory representations become less reliable over time. As shown by the authors in Figure 1c and Figure 4a, the on-diagonal decoding accuracy generally decreased over time. This implies that the signal-to-noise ratio was decreasing over time. Classifiers trained with data of relatively higher SNR and lower SNR may rely on different features, leading to poor generalization performance. This issue should be addressed in the paper.
We thank the reviewer for raising this issue and we have now run three simulations that aim to address whether a changing SNR across time might create dynamic clusters.
In the first simulation we created a dataset of 200 voxels that have a sine or cosine response function to orientations between 1° to 180°, the same orientations as the remembered target. A circular shift is applied to each voxel to vary preferred (or maximal) responses of each simulated voxel. We then assess the decoding performance under different SNR conditions during training and testing. For each of the seven iterations we selected 108 responses (out of 180) to train on and 108 to test on. To increase variability the selected trials differed in each iteration. Random white noise was applied to the data and thus the SNR was independently scaled according to the specified levels for train and test data. We then use the same pSVR decoder as in the temporal cross decoding analysis to train and test.
The second and third simulations more directly address whether increased noise levels would induce the decoder to rely on different features of the no-distractor and noise distractor data. We use empirical data from the primary visual cortex (V1; where dynamic coding was seen in the noise distractor trials) under the no-distractor and noise distractor conditions for the second and third simulations, respectively. Data from time points 5.6–8.8 seconds after stimulus onset are averaged across five TRs. As in the first simulation, SNR is systematically manipulated by adding white noise. Additionally, to see whether the initial decrease in SNR and subsequent increase would result in dynamic coding clusters, we initially increased and subsequently decreased the amplitude of added noise. The same pSVR decoder was used to train and test on the data with different levels of added noise.
We see an absence of dynamic elements in the SNR cross-decoding matrices, as the decoding accuracy primarily depends on the training data rather than test data. This results in some off-diagonal values in the decoding matrix that are higher, rather than smaller, than corresponding on-diagonal elements.
We have now added a Methods section explaining the simulations in more detail and Supplementary Figure 9 showing the SNR cross-decoding matrices.
(2) The paper tests against a strong version of stable coding, where neural spaces representing WM contents must remain identical over time. In this version, any changes in the neural space will be evidence of dynamic coding. As the paper acknowledges, there is already ample evidence arguing against this possibility. However, the evidence provided here (dynamic coding cluster, angle between coding spaces) is not as strong as what prior studies have shown for meaningful transformations in neural coding. For instance, the principal angle between coding spaces over time was smaller than 8 degrees, and around 7 degrees between sensory distractors and WM contents. This suggests that the coding space for WM was largely overlapping across time and with that for sensory distractors. Therefore, the major conclusion that working memory contents are dynamically coded is not well-supported by the presented results.
We thank the reviewer for this comment. The principal angles we calculate are above-baseline, meaning that we subtract the within-subspace principal angles from the between-subspace principal angles and take the average. Thus a 7 degree difference does not imply that there are only 7 degrees separating e.g. the sensory distractor from the target; it just indicates that the separation is 7 degrees above chance.
(3) Relatedly, the main conclusions, such as "VWM code in several visual regions did not generalize well between different time points" and "VWM and feature-matching sensory distractors are encoded in separable coding spaces" are somewhat subjective given that cross-condition generalization analyses consistently showed above chance-level performance. These results could be interpreted as evidence of stable coding. The authors should use more objective descriptions, such as 'temporal generalization decoding showed reduced decoding accuracy in off-diagonals compared to on-diagonals.
Thank you, we agree that our previous claims might have been too strong. We have now toned down our statements in the Abstract and use “did not fully generalize” and “VWM and feature-matching sensory distractors are encoded in coding spaces that do not fully overlap.”
Reviewer #2 (Recommendations For The Authors):
Weakness 1 can potentially be addressed with data simulations that fix the signal pattern, vary the noise pattern, and perform the same temporal generalization analysis to test whether changes in SNR can lead to seemingly dynamic coding formats.
Thank you for the great suggestion. We have now run the suggested simulations. Please see above (response to Weakness 1).
There are mismatches in the statistical symbols shown in Figure 4 and Supplementary Table 2. It seems that there was a swap between the symbols for the noise between-condition and noise within-condition.
Thank you, this has now been fixed.
-
-
eLife assessment
This useful study reports a reanalysis of one experiment of a previously published report to characterize the dynamics of neural population codes during visual working memory in the presence of distracting information. The evidence supporting the claims of dynamic codes is incomplete, as only a subset of the original data is analyzed, there is only modest evidence for dynamic coding in the results, and the result might be affected by the signal-to-noise ratio. This research will be of interest to cognitive neuroscientists working on the neural bases of visual perception and memory.
-
Reviewer #1 (Public Review):
Summary:
In this study, the authors re-analyzed Experiment 1 of a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly oriented grating). In this manuscript, the authors focused on identifying whether the neural code in the retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but …
Reviewer #1 (Public Review):
Summary:
In this study, the authors re-analyzed Experiment 1 of a public dataset (Rademaker et al, 2019, Nature Neuroscience) which includes fMRI and behavioral data recorded while participants held an oriented grating in visual working memory (WM) and performed a delayed recall task at the end of an extended delay period. In that experiment, participants were pre-cued on each trial as to whether there would be a distracting visual stimulus presented during the delay period (filtered noise or randomly oriented grating). In this manuscript, the authors focused on identifying whether the neural code in the retinotopic cortex for remembered orientation was 'stable' over the delay period, such that the format of the code remained the same, or whether the code was dynamic, such that information was present, but encoded in an alternative format. They identify some time points - especially towards the beginning/end of the delay - where the multivariate activation pattern fails to generalize to other time points and interpret this as evidence for a dynamic code. Additionally, the authors compare the representational format of remembered orientation in the presence vs absence of a distracting stimulus, averaged over the delay period. This analysis suggested a 'rotation' of the representational subspace between distracting orientations and remembered orientations, which may help preserve simultaneous representations of both remembered and viewed stimuli.
Strengths:
(1) Direct comparisons of coding subspaces/manifolds between time points and task conditions is an innovative and useful approach for understanding how neural representations are transformed to support cognition.
(2) Re-use of existing datasets substantially goes beyond the authors' previous findings by comparing the geometry of representational spaces between conditions and time points, and by looking explicitly for dynamic neural representations
Weaknesses:
(1) Only Experiment 1 of Rademaker et al (2019) is reanalyzed. The previous study included another experiment (Expt 2) using different types of distractors which did result in distractor-related costs to neural and behavioral measures of working memory. The Rademaker et al (2019) study uses these two results to conclude that neural WM representations are protected from distraction when distraction does not impact behavior, but conditions that do impact behavior also impact neural WM representations. Considering this previous result is critical for relating the present manuscript's results to the previous findings, it seems necessary to address Experimentt 2's data in the present work
(2) Primary evidence for 'dynamic coding', especially in the early visual cortex, appears to be related to the transition between encoding/maintenance and maintenance/recall, but the delay period representations seem overall stable, consistent with previous findings
(3) Dynamicism index used in Figure 1f quantifies the proportion of off-diagonal cells with significant differences in decoding performance from the diagonal cell. It's unclear why the proportion of time points is the best metric, rather than something like a change in decoding accuracy. This is addressed in the subsequent analysis considering coding subspaces, but the utility of the Figure 1f analysis remains weakly justified.
(4) There is no report of how much total variance is explained by the two PCs defining the subspaces of interest in each condition, and timepoint. It could be the case that the first two principal components in one condition (e.g., sensory distractor) explain less variance than the first two principal components of another condition.
(5) Converting a continuous decoding metric (angular error) to "% decoding accuracy" serves to obfuscate the units of the actual results. Decoding precision (e.g., sd of decoding error histogram) would be more interpretable and better related to both the previous study and behavioral measures of WM performance.
(6) This report does not make use of behavioral performance data in the Rademaker et al (2019) dataset.
(7) Given there were observed differences between individual retinotopic ROIs in the temporal cross-decoding analyses shown in Figure 1, the lack of data presented for the subspace analyses for the corresponding individual ROIs is a weakness
-
Reviewer #2 (Public Review):
Summary:
In this work, Degutis and colleagues addressed an interesting issue related to the concurrent coding of sensory percepts and visual working memory contents in visual cortices. They used generalization analyses to test whether working memory representations change over time, diverge from sensory percepts, and vary across distraction conditions. Temporal generalization analysis demonstrated that off-diagonal decoding accuracies were lower than on-diagonal decoding accuracies, regardless of the presence of intervening distractions, implying that working memory representations can change over time. They further showed that the coding space for working memory contents showed subtle but statistically significant changes over time, potentially explaining the impaired off-diagonal decoding performance. The …
Reviewer #2 (Public Review):
Summary:
In this work, Degutis and colleagues addressed an interesting issue related to the concurrent coding of sensory percepts and visual working memory contents in visual cortices. They used generalization analyses to test whether working memory representations change over time, diverge from sensory percepts, and vary across distraction conditions. Temporal generalization analysis demonstrated that off-diagonal decoding accuracies were lower than on-diagonal decoding accuracies, regardless of the presence of intervening distractions, implying that working memory representations can change over time. They further showed that the coding space for working memory contents showed subtle but statistically significant changes over time, potentially explaining the impaired off-diagonal decoding performance. The neural coding of sensory distractions instead remained largely stable. Generalization analyses between target and distractor codes showed overlaps but were not identical. Cross-condition decodings had lower accuracies compared to within-condition decodings. Finally, within-condition decoding revealed more reliable working memory representations in the condition with intervening random noises compared to cross-condition decoding using a trained classifier on data from the no-distraction condition, indicating a change in the VWM format between the noise distractor and no-distractor trials.
Strengths:
This paper demonstrates a clever use of generalization analysis to show changes in the neural codes of working memory contents across time and distraction conditions. It provides some insights into the differences between representations of working memory and sensory percepts, and how they can potentially coexist in overlapping brain regions.
Weaknesses:
(1) An alternative interpretation of the temporal dynamic pattern is that working memory representations become less reliable over time. As shown by the authors in Figure 1c and Figure 4a, the on-diagonal decoding accuracy generally decreased over time. This implies that the signal-to-noise ratio was decreasing over time. Classifiers trained with data of relatively higher SNR and lower SNR may rely on different features, leading to poor generalization performance. This issue should be addressed in the paper.
(2) The paper tests against a strong version of stable coding, where neural spaces representing WM contents must remain identical over time. In this version, any changes in the neural space will be evidence of dynamic coding. As the paper acknowledges, there is already ample evidence arguing against this possibility. However, the evidence provided here (dynamic coding cluster, angle between coding spaces) is not as strong as what prior studies have shown for meaningful transformations in neural coding. For instance, the principal angle between coding spaces over time was smaller than 8 degrees, and around 7 degrees between sensory distractors and WM contents. This suggests that the coding space for WM was largely overlapping across time and with that for sensory distractors. Therefore, the major conclusion that working memory contents are dynamically coded is not well-supported by the presented results.
(3) Relatedly, the main conclusions, such as "VWM code in several visual regions did not generalize well between different time points" and "VWM and feature-matching sensory distractors are encoded in separable coding spaces" are somewhat subjective given that cross-condition generalization analyses consistently showed above chance-level performance. These results could be interpreted as evidence of stable coding. The authors should use more objective descriptions, such as 'temporal generalization decoding showed reduced decoding accuracy in off-diagonals compared to on-diagonals.
-
