Testosterone-induced metabolic changes in seminal vesicle epithelium modify seminal plasma components with potential to improve sperm motility
Curation statements for this article:-
Curated by eLife

eLife Assessment
This important work elucidates the biological processes and detailed mechanisms by which testosterone influences seminal plasma metabolites in mice. The evidence supporting the upregulation of metabolic enzymes and the role of ACLY is solid, highlighting the potential contributions of fatty acids to sperm motility.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- @yangbb's saved articles (yangbb)
- Evaluated articles (eLife)
Abstract
Male factors account for almost half of the causes of infertility. In rodents and humans, most of the components of semen are supplied by the seminal vesicles, and they support male reproductive ability, but there are many unknown details. This study focused on metabolic changes in seminal vesicle epithelial cells and investigated how testosterone affects seminal plasma composition. A factor improving the linear motility of sperm was secreted from the seminal vesicles, and it was produced in an androgen-dependent manner. Bioassays, gene expression, and flux analysis studies demonstrated that testosterone promotes glucose uptake in seminal vesicle epithelial cells via GLUT4, resulting in fatty acid synthesis. ACLY was a critical factor in this metabolic change, which produced fatty acids, especially oleic acid. In conclusion, the critical role of testosterone-induced metabolic changes in the seminal vesicles is to ensure the synthesis of fatty acids. These findings suggest that testosterone-dependent lipid remodeling may contribute to sperm straight-line motility, and further functional verification is required.
Article activity feed
-

-
-
-

eLife Assessment
This important work elucidates the biological processes and detailed mechanisms by which testosterone influences seminal plasma metabolites in mice. The evidence supporting the upregulation of metabolic enzymes and the role of ACLY is solid, highlighting the potential contributions of fatty acids to sperm motility.
-
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors have made improvements from the previous version by softening the claim that oleic acid derived from seminal vesicle epithelium strongly affects linear progressive motility in isolated cauda epididymal sperm in vitro. They have also addressed the ambiguous references to the strength of the relationship between fatty acids and sperm motility, making the manuscript more balanced and nuanced.
Strengths:
This study addresses an important gap in our understanding of how testosterone influences seminal plasma metabolites and, in turn, sperm motility. The findings provide valuable insights into the sensitivity of seminal vesicle epithelial cells to …
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors have made improvements from the previous version by softening the claim that oleic acid derived from seminal vesicle epithelium strongly affects linear progressive motility in isolated cauda epididymal sperm in vitro. They have also addressed the ambiguous references to the strength of the relationship between fatty acids and sperm motility, making the manuscript more balanced and nuanced.
Strengths:
This study addresses an important gap in our understanding of how testosterone influences seminal plasma metabolites and, in turn, sperm motility. The findings provide valuable insights into the sensitivity of seminal vesicle epithelial cells to testosterone, which could improve in vitro conditions for studying sperm motility. The authors have added methodological details and re-performed experiments with more appropriate control groups, enhancing the robustness of the study. These revisions, along with more carefully modified language reflecting measurement nuances, add significant value to the field. The study's detailed exploration of the physiological role of reproductive tract glandular secretions in modulating sperm behaviors is likely to be of broad interest, providing a strong foundation for future research on the relationship between fatty acid beta-oxidation and sperm motility patterns.
Weaknesses:
While the connection between media fatty acids and sperm motility patterns is still not fully conclusive, the authors have taken substantial steps to clarify and tone down their conclusions. The revised manuscript presents a more balanced view, acknowledging the complexity of the relationship and providing a more solid basis for follow-on studies.
-
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels, as well as isolated mouse and human seminal vesicle epithelial cells, the authors demonstrate that testosterone induces an increase in glucose uptake. The study reveals that testosterone triggers differential gene expression, particularly focusing on metabolic enzymes. They specifically identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to heightened production of 18:1 oleic acid. The revised version of the manuscript significantly strengthens the role of ACLY as a central regulator of seminal vesicle epithelial cell metabolic programming. The authors suggest that fatty acids secreted by seminal vesicle epithelial cells are taken up by sperm, …
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels, as well as isolated mouse and human seminal vesicle epithelial cells, the authors demonstrate that testosterone induces an increase in glucose uptake. The study reveals that testosterone triggers differential gene expression, particularly focusing on metabolic enzymes. They specifically identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to heightened production of 18:1 oleic acid. The revised version of the manuscript significantly strengthens the role of ACLY as a central regulator of seminal vesicle epithelial cell metabolic programming. The authors suggest that fatty acids secreted by seminal vesicle epithelial cells are taken up by sperm, resulting in a positive impact on sperm function. While the lipid mixture mimicking the lipids secreted by seminal vesicle epithelial cells shows marginal positive effect on sperm motility, the authors have made considerable progress in refining their conclusions. The revised manuscript acknowledges the complexity of pinpointing the specific seminal vesicle fluid component that potentially positively affects sperm function, providing a more measured and credible interpretation of their findings.
-
Author response:
The following is the authors’ response to the previous reviews
Editor's note:
Thank you for taking time and efforts to improve this study. After re-review, two reviewers have a consensus that the connections the fatty acids and sperm motility is still ambiguous. Thus, I recommend to further tone down this conclusion consistently in the title and the text pointed out by reviewers before making a final version of record.
We sincerely appreciate the considerable time and effort you and the reviewers devoted to evaluating our manuscript. We have revised the title and text to express the relationship between fatty acids and sperm motility more consistently and toned down. With these revisions, we would like to proceed with publishing the manuscript as the Version of Record (VoR). Thank you very much for your guidance in …
Author response:
The following is the authors’ response to the previous reviews
Editor's note:
Thank you for taking time and efforts to improve this study. After re-review, two reviewers have a consensus that the connections the fatty acids and sperm motility is still ambiguous. Thus, I recommend to further tone down this conclusion consistently in the title and the text pointed out by reviewers before making a final version of record.
We sincerely appreciate the considerable time and effort you and the reviewers devoted to evaluating our manuscript. We have revised the title and text to express the relationship between fatty acids and sperm motility more consistently and toned down. With these revisions, we would like to proceed with publishing the manuscript as the Version of Record (VoR). Thank you very much for your guidance in improving our study.
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. Based on limited evidence in previous versions of the report, the authors softened the claim that oleic acid derived from seminal vesicle epithelium strongly affects linear progressive motility in isolated cauda epididymal sperm in vitro. Though the report still contains somewhat ambiguous references to the strength of the relationship between fatty acids and sperm motility.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as add important physiological context to the role of reproductive tract glandular secretions in modulating sperm behaviors. The strongest observations are related to the sensitivity of seminal vesicle epithelial cells to testosterone. The revisions include the addition of methodological detail, modified language to reflect the nuance of some of the measurements, as well as re-performed experiments with more appropriate control groups. The findings are likely to be of general interest to the field by providing context for follow-on studies regarding the relationship between fatty acid beta oxidation and sperm motility pattern.
Weaknesses:
The connection between media fatty acids and sperm motility pattern remains inconclusive.
We would like to express our sincere gratitude to the judges for their cooperation in reviewing the manuscript and for your helpful comments, which were instrumental in improving manuscript.
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. The authors propose that fatty acids are secreted by seminal vesicle epithelial cells and are taken up by sperm, positively affecting sperm function. A lipid mixture mimicking the lipids secreted by seminal vesicle epithelial cells, however, only has a small and mostly non-significant effect on sperm motility, suggesting the authors were not apply to pinpoint the seminal vesicle fluid component that positively affects sperm function.
We greatly appreciate the reviewer’s thoughtful comments and time spent reviewing this manuscript. The relationship between lipids such as fatty acids and sperm motility remains unclear in the current dataset. Therefore, before finalizing the manuscript, we revised the title and text, as suggested by the reviewers, to express this conclusion more cautiously and consistently.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Some additional comments are provided below to aid the authors in improving the quality of the work:
Major Comments:
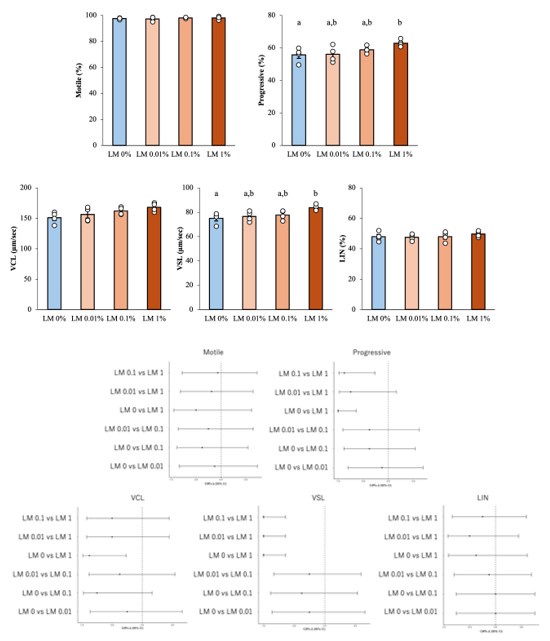
(1) In the newly added supplemental figure 5, the authors note that the percentage data were arcisine transformed prior to statistical analysis without providing any other justification. This seems strange, especially for such a small sample size. It seems more appropriate for the authors to use a nonparametric test. Forcing symmetry without knowing what the shape of the true distribution is makes the ANOVA hard to interpret. Additionally, why use pairwise comparisons rather than comparing each group to the control (LM 0%). Also, note that the graphs are not individually labeled to distinguish them in the legend (A, B, C, etc.). Ultimately, the treatment differences don't seem that meaningful, even if the authors were able to observe statistical significance with the somewhat over-manipulated method of analysis.
Ultimately, the conclusion of this experiment (Supplemental figure 5) remains unchanged, but we agree that the relationship between fatty acids and sperm motility remains unclear. Therefore, before finalizing the manuscript, we revised the title and text as pointed out by the reviewers to express this conclusion more cautiously and consistently throughout the manuscript.
Arcsin transform is commonly used for percentage data [Zar, J.H. 2010. Biostatistical analysis., McDonald, J.H. 2014. Handbook of biological statistics.]. If the values are low or high, such as 0 to 30% or 70 to 100%, without arcsine transformation will result in a large deviation from the normality of the data. However, even if such a conversion is performed, it does not necessarily mean that the assumptions of normality and homogeneity of variance, which are prerequisites for parametric statistical analysis methods, are satisfied.
Given the small sample size and the possibility of non-normal data, we performed Shapiro–Wilk tests for each group (n = 6) and found no departure from normality (all p > 0.1). Q–Q plots and Levene’s test (p > 0.1) likewise supported the assumptions of ANOVA. Following the reviewer’s recommendation, we repeated the analysis with a Kruskal–Wallis test followed by Dunn’s post-hoc comparisons (Bonferroni corrected). Both approaches led to the same conclusions, with non-parametric p-values equal to or smaller than the parametric ones. In the revised manuscript we now report ANOVA as the primary analysis. The author response image includes effect sizes with 95 % confidence intervals, and provide the non-parametric results for transparency.
Author response image 1.
Results of reanalysis of supplementary Figure 5 using nonparametric tests and effect sizes with 95% confidence intervals. Upper part; Differences between groups were assessed by Kruskal–Wallis test, differences among values were analyzed by Dunn’s post-hoc comparisons (Bonferroni corrected) for multiple comparisons. Different letters represent significantly different groups. Lower part; The effect sizes with 95 % confidence intervals. For example, Cliff's Δ = -1 (95% CI ~ -0.6) in VSL's “LM 0 vs LM1” means that LM 1% values exceed LM 0 %values in all pairs.
(2) I appreciate that the authors toned down the interpretation of the effects of seminal plasma metabolites on sperm motility with a cautionary statement on Lines 397-405 and Line 259. However, they send mixed signals with the title of the report: "Testosterone-Induced Metabolic Changes in Seminal Vesicle Epithelial cells Alter Plasma Components to Enhance Sperm Motility", and on line 265 when the say "ACLY expression is upregulated by testosterone and is essential for the metabolic shift of seminal vesicle epithelial cells that mediates sperm linear motility".
The wording has been softened overall. The title has been changed to “Testosterone-Induced Metabolic Changes in Seminal Vesicle Epithelium Modify Seminal Plasma Components with Potential to Improve Sperm Motility” In the results (lines 265-266), we have stated that “ACLY expression is upregulated by testosterone and is essential for the metabolic shift that is associated with increased linear motility” without implying a causal relationship.
Minor Comments:
(1) Typo on line 31: "understanding the male fertility mechanisms and may perspective for the development of potential biomarkers of male fertility and advance in the treatment of male infertility."
We have made the following corrections. “These findings suggest that testosterone-dependent lipid remodeling may contribute to sperm straight-line motility, and further functional verification is required.”
(2) Line 193: the statement is confusing "Therefore, we analyzed mitochondrial metabolism using a flux analyzer, predicting that more glucose is metabolized, pyruvate is metabolized from phosphoenolpyruvic acid through glycolysis in response to testosterone, and is further metabolized in the mitochondria." For example, 'Metabolized through glycolysis' is an ambiguous way to describe the pyruvate kinase reaction. Additionally, phosphoenolpyruvate has three acid ionizable groups, two of which have pKa's well below physiological pH, so phosphoenolpyruvate is the correct intermediate rather than phosphoenolpyruvic acid. The authors make similar mistakes with other organic acids such as citric acid.
Rewritten as “We therefore examined cellular energy metabolism with a flux analyzer, anticipating that testosterone would elevate glycolytic flux, thereby producing more pyruvate from phosphoenolpyruvate. Because extracellular pyruvate levels simultaneously declined, we inferred that the cells had an increased pyruvate demand and, at that time, hypothesized that the excess pyruvate would enter the mitochondria to support enhanced oxidative metabolism.” (lines 193-198)
The organic acids are now referenced in their appropriate forms (e.g., citrate, phosphoenolpyruvate).
(3) Line: 271: "Acly" should be all capitalized to "ACLY". The report mixes capitalizing through out and could be more consistent.
We appreciate the reviewer’s attention to nomenclature and have standardized the manuscript accordingly. Proteins are written in Roman letters, all in capital letters. Mouse gene symbols: italics, first letter capitalize.
Reviewer #2 (Recommendations for the authors):
Major comments:
(1) 'Once capacitation is complete, sperm cannot maintain that state for a long time'. The publications cited by the author do not support that statement and this reviewer also does not agree. Lower fertilization efficiency from in vitro capacitated epidydimal sperm does not have to mean capacitation is reversed, it can simply mean in vitro capacitation conditions not accurately mimic capacitation in vivo.
We thank the reviewer for pointing this out and would like to clarify our position. Our statement does not suggest a "reversal" of active capacitation. Rather, it reflects the well-documented fact that capacitation is a transient process. Sperm that undergo capacitation too early cannot maintain that state for long enough to retain their ability to fertilize at the moment and location of fertilization in vivo.
(2) How do the authors explain the discrepancy between the results shown in Fig. S1E, the increase in sperm motility upon mixing of sperm with SVF and the results reported in Li et al 2025. Mentioning decapacitating factors without further explanation is insufficient.
We appreciate the reviewer's feedback pointing out the need for a clearer explanation.
Seminal plasma is inherently binary, containing both decapacitation factors that delay or inhibit capacitation and nutrient substrates that promote sperm motility.
In vivo, it is believed that the coating of sperm by decapacitation factors is removed by uterine fluid and albumin as it passes through the female reproductive tract [PMID: 22827391, PMID: 24274412]. In contrast, standard fertilization culture media lack a clearance pathway, so decapacitating factors are retained throughout the culture period. As a result, the cleavage rate after in vitro fertilization using sperm exposed to seminal vesicle fluid decreased dramatically.
Lipids, such as fatty acids, increased sperm motility without directly inducing markers of fertilization. These results suggest that the enhancement of motility by lipids is functionally distinct from the capacitation-inhibiting function of seminal plasma proteins. The data from this study are consistent with the biphasic model. Specifically, decapacitation factors temporarily stabilize the sperm membrane, preventing early capacitation. Meanwhile, lipids enhance sperm motility, enabling them to rapidly pass through the hostile uterine environment.
(3) This reviewer does not see the merit in including a lipid mixture motility experiment compared to using OA alone. The increase in motility is still small and far from comparable to the motility increase with seminal vesicle fluid. In this reviewer's opinion the experiment is still inconclusive and should not be highlighted in the manuscript title.
The wording has been softened overall. The title has been changed to “Testosterone-Induced Metabolic Changes in Seminal Vesicle Epithelium Modify Seminal Plasma Components with Potential to Improve Sperm Motility”. (Please see also Reviewer 1's main comment 1)
Minor comments:
(1) 'This change includes a large amplitude of flagella' does not make sense. Please correct.
The following corrections have been made. “This change is characterized by large-amplitude flagellar beating.” (lines 44-45)
-
-
eLife Assessment
This important work shows the biological processes and detailed mechanisms involving testosterone's influence on seminal plasma metabolites in mice. Evidence supporting the up regulation of metabolic enzymes and the role of ACLY is solid, though the precise contributions of fatty acids to sperm motility requires further elucidation.
-
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. Based on limited evidence in previous versions of the report, the authors softened the claim that oleic acid derived from seminal vesicle epithelium strongly affects linear progressive motility in isolated cauda epididymal sperm in vitro. Though the report still contains somewhat ambiguous references to the strength of the relationship between fatty acids and sperm motility.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as …
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. Based on limited evidence in previous versions of the report, the authors softened the claim that oleic acid derived from seminal vesicle epithelium strongly affects linear progressive motility in isolated cauda epididymal sperm in vitro. Though the report still contains somewhat ambiguous references to the strength of the relationship between fatty acids and sperm motility.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as add important physiological context to the role of reproductive tract glandular secretions in modulating sperm behaviors. The strongest observations are related to the sensitivity of seminal vesicle epithelial cells to testosterone. The revisions include the addition of methodological detail, modified language to reflect the nuance of some of the measurements, as well as re-performed experiments with more appropriate control groups. The findings are likely to be of general interest to the field by providing context for follow-on studies regarding the relationship between fatty acid beta oxidation and sperm motility pattern.
Weaknesses:
The connection between media fatty acids and sperm motility pattern remains inconclusive.
-
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. The authors propose that fatty acids are secreted by seminal vesicle epithelial cells and are taken up by sperm, positively affecting sperm function. A lipid mixture …
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. The authors propose that fatty acids are secreted by seminal vesicle epithelial cells and are taken up by sperm, positively affecting sperm function. A lipid mixture mimicking the lipids secreted by seminal vesicle epithelial cells, however, only has a small and mostly non-significant effect on sperm motility, suggesting the authors were not apply to pinpoint the seminal vesicle fluid component that positively affects sperm function.
-
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors identify oleic acid as a particularly important metabolite, derived from seminal vesicle epithelium, that stimulates linear progressive motility in isolated cauda epidydimal sperm in vitro. The authors provide additional experimental evidence of a testosterone dependent mechanism of oleic acid production by the seminal vesicle epithelium.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this …
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors identify oleic acid as a particularly important metabolite, derived from seminal vesicle epithelium, that stimulates linear progressive motility in isolated cauda epidydimal sperm in vitro. The authors provide additional experimental evidence of a testosterone dependent mechanism of oleic acid production by the seminal vesicle epithelium.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as add important physiological context to the role of reproductive tract glandular secretions in modulating sperm behaviors. The strongest observations are related to the sensitivity of seminal vesicle epithelial cells to testosterone. The revisions include addition of methodological detail, modified language to reflect the nuance of some of the measurements, as well as re-performed experiments with more appropriate control groups. The findings are likely to be of general interest to the field by providing context for follow-on studies regarding the relationship between fatty acid beta oxidation and sperm motility pattern.
Thank you for summarizing and your positive evaluation of our study.
Weaknesses:
Support for the proposed mechanism is stronger in this revised report than in the previous report, but there are many challenges in measuring sperm metabolism and its direct relationship with motility patterns. This study is no exception and largely relies on correlations between various experiments in lieu of direct testing. Additionally, the discussion is framed from a human pre-clinical perspective, and it should be noted that the reproductive physiology between mice and humans is very different.
Thank you for pointing out the challenges in our paper. We appreciate your comment on the limited evidence supporting the direct relationship between sperm metabolism and motility patterns under current experimental conditions. Based on your and reviewer2’s suggestions, we have decided to remove the experiments and discussion on the “effects of OA on sperm metabolism, motility and fertility (Fig. 7, Supplemental Figure 5A and C-F.)” and the corresponding parts in the Discussion section from the paper. (See also Reviewer 2's main comment) These data mainly show correlations, and did not show direct evidence of causality. Instead, we added a new experiment to the manuscript, in which a lipid mixture that mimics the fatty acid profile secreted testosterone-dependently from seminal vesicle epithelial cells was added to the sperm culture medium (New Supplemental Figure 5, Lines 259-268). In this experiment, motility parameters were measured using CASA. This experiment evaluates the direct effects of lipid exposure on sperm motility. With these revisions, we are able to focus on the metabolic changes caused by testosterone in seminal vesicle epithelial cells, which are the central focus of our research. We have added a short statement agreeing the potential importance of OA and our intention to more rigorously investigate the role of OA in sperm function in subsequent studies (Lines 402-407).
Furthermore, we have revised text, clearly state the limitations of the species difference and clarify that the translational aspects to humans are speculative (Lines 383-384, 395-397, 408-410).
We appreciate your guidance. We believe that these changes will strengthen our research.
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. 18:1 oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility and in vivo fertilization in the presence of 18:1 oleic acid and the absence of testosterone, however, is small. Additional experiments should be included to further support that oleic acid positively affects sperm function.
Thank you very much for carefully reading the manuscript and for your comments. We appreciate your understanding that the role of ACLY in metabolic programming of seminal vesicle epithelial cells has been strengthened in the revised version. On the other hand, we agree with your view that the increase in sperm motility and fertilization rate by oleic acid is minimal under the current experimental conditions. We agree that further evidence is needed to support our conclusion regarding the positive effects of oleic acid on sperm function. Based on your comments and our re-evaluation of the data, we have decided to remove the experiments and discussion on “OA and sperm motility” from the current paper (Fig. 7, Supplemental Figure 5A and C-F). In the revised paper, we have significantly toned down the claims on the previous role of oleic acid and instead focused on the metabolic regulatory mechanisms of seminal vesicle epithelial cells.
We hope that these revisions address your concerns and improve the overall clarity of the manuscript.
Recommendations for the authors:
Note from the reviewing editor: The reviewers agree that the revised manuscript is significantly improved and view the work as important. Both reviewers agree that the evidence for testosterone effects on seminal vesicle epithelial cells to support fatty acid synthesis is strong and suggest that the authors tone down their conclusion of oleic acid effect on sperm motility as the effect is very small. With this minor changes, the evidence to support the conclusion of the study is viewed as solid.
Thank you for recognizing the improvements that we have made to our manuscript and for appreciating the importance of our research. We also appreciate your assessment that the evidence for the effect of testosterone on seminal vesicle epithelial cells that support fatty acid synthesis is solid.
On the other hand, we agree with the two reviewers that the effect of oleic acid on sperm motility is limited and that the relevant data do not measure a direct relationship. Therefore, we have decided to withdraw the data set on the effect of oleic acid on sperm (Fig. 7, Supplemental Figure 5A and C-F) and focus this paper on seminal vesicle epithelial cells (in response to reviewer 2's suggestion). Given that testosterone-induced lipid (Fatty acid) synthesis in seminal vesicle epithelial cells is a key aspect of our study, we have included additional experiments in the revised manuscript to show how lipids affect sperm (New Supplemental Figure5, Lines 259-263).
With these revisions, the manuscript emphasizes the importance of testosterone-dependent fatty acid synthesis in seminal vesicle epithelial cells and the fact that this includes oleic acid. The title has also been partially revised in line with these revisions.
Reviewer #1 (Recommendations for the authors):
Minor Comments:
(1) The authors indicate in the methods that extracellular flux analysis was normalized to cell count. However, the y-axis units in Figs 4, 8, 9 and SFig 9 are not normalized.
(2) The OA label appears to be missing from Fig 7A. Additionally, the scale bar is offset in one of the images and the length of the scale bar does not appear to be mentioned in the figure legend.
Thank you for raising these points. We have corrected.
Fig. 7 has been withdrawn in response to Reviewer 2's suggestion.
Reviewer #2 (Recommendations for the authors):
With the experiments included in their revised version the authors strengthen their conclusions about testosterone-induced metabolic reprogramming in seminal vesicle cells resulting in reduced proliferation. The experiments surrounding ACLY are well-designed and give insights into the underlying molecular mechanisms. For other parts, the manuscript became less clear and it is often hard to follow the author's line of thoughts for their conclusions.
Based on the experiments shown in the manuscript this reviewer is still not convinced that OA positively affects sperm function. The changes in linear motility are minor, blastocyst levels are lower and the authors do not show that OA alone positively affects cleavage rate during AI. Without additional experiments that show a stronger effect on sperm function, the authors should consider focusing the manuscript exclusively on seminal vesicle epithelial cells.
Thank you for your constructive comments on our paper. We thank the reviewer for pointing out that the effect of oleic acid (OA) on sperm function is limited in our current experiments. As reviewer 1 also pointed out, we agree that further experiments and improved methodology are needed to reliably demonstrate the functional effects of OA on sperm. Because the strength of the data on the direct relationship between fatty acids in seminal fluid and improved sperm function is currently insufficient, we have removed the data set for oleic acid and sperm motility (Fig. 7, Supplemental Figure 5A and C-F) and focused on the “the mechanism of metabolic regulation of testosterone in seminal vesicle epithelial cells”. We have consistently narrowed the focus of the paper to the theme of “how testosterone changes energy metabolism in seminal vesicle epithelial cells”. In accordance with this change, the structure of the paper has also been partially revised (red text in the manuscript). With these revisions, the main point of the paper focuses on the mechanism by which testosterone regulates metabolic pathways in the seminal vesicle epithelial cells.
For more detailed revisions, please see the responses to your comments below.
(1) 45-55 still need major revision. It will not become clear to the reader what the authors mean by epididymal maturation. 'Ability to fertilize in in vitro?' Epididymal sperm are moving linearly in the absence of seminal vesicle fluid. Increased progressive motility, hyperactivation, and the ability to undergo the acrosome reaction are induced upon exposure to seminal vesicle fluid. The authors should introduce the concept of capacitation and that capacitation can be induced in vitro by exposure to bicarbonate and a cholesterol acceptor.
Thank you for pointing out the ambiguity of epididymal maturation, the need to clarify the concept of capacitation, and the role of seminal plasma in this context. The revised text explains that epididymal maturation only gives sperm their potential ability to fertilize. It also explains that it is the subsequent capacitation process—inducible in vitro by incubation with bicarbonate and cholesterol acceptors—that gives full fertilization potential. On the other hands, we emphasize that in vivo, seminal plasma, which contains both capacitation-promoting and decapacitation factors, plays a key role in fine-tuning the timing of capacitation, ensuring that sperm acquire fertilization competence at the appropriate moment. We hope that these revisions clarify our intended meaning and strengthen the overall message of the paragraph. (lines 42-54)
“Sperm that have completed spermatogenesis in the testis acquire their potential to fertilize while maturing in the epididymis (5–7). The physiological change of sperm during fertilization process are collectively referred to as “capacitation”. This change includes a large amplitude of flagella (called hyperactivation) and developing the capacity to undergo the acrosome reaction, and can be induced by culturing sperm collected from the epididymis in a medium containing bicarbonate and cholesterol acceptors (8, 9). However, once capacitation is complete, sperm cannot maintain that state for a long time. Therefore, even if epididymal sperm that have not been exposed to seminal plasma are artificially inseminated into the cervix or uterus, the fertilization rate remains low (10–12). That is because, in vivo, during ejaculation, exposure of epididymal sperm to seminal plasma masks the unintended capacitation as they pass through the female reproductive tract and ensures fertilization of sperm that reach the oviduct (13). In other words, seminal plasma plays an important role in fine-tuning the timing of sperm capacitation and in maintaining the sustained sperm motility needed to reach the oviduct.”
(2) 81: Similar as in their rebuttal the authors should further elute on the connection between fructose, citrate, and testosterone. That still does not become clear. Based on the author's explanation in the rebuttal, why are citrate and fructose levels higher when the animals are castrated?
We thank you for the opportunity to clarify our statement regarding the relationship between fructose, citrate, and testosterone. Our original explanation was intended to reflect the fact that testosterone from the testes has a stimulating effect on the accessory reproductive glands, and to report that the concentrations of fructose and citric acid were higher in the non-castrated (control) animals than in the castrated animals. In castrated animals, the absence of testosterone leads to decreased activity of these glands and, consequently, lower levels of these metabolites. To make this clear, we have revised the manuscript as follows. (lines 76-82)
“Several specific factors produced by the male accessory glands that contribute to seminal plasma and impact male fertility have been elucidated. For example, surgical removal of seminal vesicles in male mice and rats was associated with infertility (17, 22, 23). The observations that fructose (24) and citric acid (25) concentrations in seminal plasma of control mice and rats are higher than in castrated animals suggest that the specific metabolism of the accessory glands might be affected by testosterone derived from the testes, which activate intracellular androgen receptors (AR; NR3C4) required for gene regulation of transcription.”
(3) 111: This reviewer does not understand the author's obsession with reporting linear motility. Sperm are moving linearly when isolated from the epididymis. Again, increase of progressive motility is a well-defined hallmark of capacitation and primarily used in the field when discussing changes in sperm motility during capacitation. This reviewer is assuming that the changes in progressive vs linear motility in Fig. 7 are not significant because the data is more scattered. The % increase seems to be approximately the same. The same is true for Fig. 8. The increase in LIN is so small and not dose-dependent that this reviewer is not comfortable making that one of the main conclusions of the manuscript.
Our claim is based on the observation that seminal vesicle secretions significantly improve the linear motility (VSL and LIN) of sperm even in an environment that does not contain capacitation-inducing factors such as BSA. We interpret this as a survival strategy for sperm to pass through the female reproductive tract efficiently. Therefore, we believe that this does not mean that the meaning of “progressive motility” in the context of conventional capacitation is the same as that of progressive motility observed in seminal plasma.
However, the reviewer's point that the current data set does not sufficiently support what the minor increase in linear motility caused by oleic acid means is agreed with. Therefore, we have decided to withdraw the dataset on the effect of oleic acid on sperm motility (Fig. 7, Supplemental Figure 5A and C-F) and have revised the conclusion. (Lines 406-410)
(4) 128: For the mitochondrial membrane potential measurements the authors should mention that they included antimycin as a control. The manuscript would benefit from including scatter plots with unloaded controls to support their gating strategy. In its current stage, the gating between low and high membrane potential seems arbitrary.
Thank you for pointing this out. We have included an explanation of antimycin as a control in the main text (Lines 920-921). In addition, we have added some reference scatter plots and also added an explanation of the gating strategy between low and high membrane potentials (Supplemental Figure 1C and D, Lines 1101-1104). We hope this change will make the manuscript clearer.
(5) 190: What do the authors mean by: 'However, there was no difference in the Oligomycin-sensitive ECAR, indicating that testosterone may increase glucose metabolism but does not enhance the expression of a group of enzymes involved in the glycolytic pathway.'
Our original intention was to state that testosterone probably increases basal glycolytic flux via increased glucose uptake (as supported by the GLUT4 translocation data), but does not increase maximal glycolytic capacity, as indicated by the lack of difference in oligomycin-sensitive ECAR.
However, as Reviewer 1 previously pointed out, we agree that the assay conditions themselves, such as the use of oligomycin to inhibit oxidative mitochondria, may create non-physiological conditions and not fully reflect the energy distribution in vivo. Under these conditions, there is a possibility that the flow of glycolysis will increase artificially as a compensatory reaction, and parameters such as “maximum glycolytic capacity” should have been interpreted with caution.
Therefore, we have revised the manuscript to clarify that our data are a single-time point under defined experimental conditions and do not necessarily provide direct insight into changes in expression or activity of individual glycolytic enzymes.
“These data indicate that testosterone enhances glucose utilization. This leads to the interpretation that testosterone increases the flow of glycolysis by increasing glucose uptake and alters metabolic flux distribution.” (Lines 186-188)
(6) 205: Could the authors elaborate further on how they came to this conclusion: 'These results suggest that testosterone does not reduce transient enzyme activity in mitochondria but rather weakens the metabolic pathway of the mitochondrial TCA cycle and/or the electron transport chain due to the changes in gene expression patterns in seminal vesicle epithelial cells.' Based on their results at this point the authors have no insights about changes in enzyme activity or gene expression that might explain the phenotype.
Our statement is based on the following observations. In testosterone-treated cells, the addition of glucose increased ECAR, suggesting an increase in glycolytic flux due to an increase in glucose uptake. On the other hand, mitochondrial respiratory parameters (basal respiration, oligomycin-sensitive respiration, FCCP-uncoupled respiration, and reserve respiratory capacity) were significantly decreased under testosterone treatment.
From these results, it was speculated that testosterone promotes the redistribution of metabolic flux, directing it away from mitochondrial oxidative phosphorylation and towards the glycolytic pathway and, possibly, lipid synthesis. However, as the reviewers correctly point out, at this point, we have not directly measured changes in the activity or expression of individual enzymes in the TCA cycle or ETC. Therefore, in the next experiment, we extracted mRNA from the cells and performed gene expression analysis using real-time PCR. To make this clear, we have revised the manuscript as follows.
“Overall, these data indicate that testosterone promotes the redistribution of metabolic flux. In other words, testosterone increased glycolysis in seminal vesicle epithelial cells while decreasing mitochondrial respiration. To determine whether these changes were accompanied by changes in gene expression of specific metabolic-related enzymes, we analyzed gene expression levels.” (Lines 201-205)
(7) 219: Characterizing ACLY as an enzyme of the ETC is misleading. ACLY is a cytosolic enzyme that connects the TCA cycle with fatty acid synthesis.
We would like to thank you for pointing out that the description of the function of ACLY could be misunderstood. We agree that characterizing ACLY as an enzyme of the ETC could be misleading. Therefore, we have revised the sentence to clearly indicate that ACLY is a cytosolic enzyme that links the TCA cycle with fatty acid synthesis. The revised text is as follows:
"Interestingly, testosterone significantly increased the expression of Acly, which encodes a cytoplasmic enzyme that converts citrate transported from the TCA cycle into acetyl-CoA, a substrate that is essential for fatty acid synthesis." (lines216-218)
(8) 228: Which results support that ETC proteins were upregulated by flutamide?
We appreciate the reviewer for this point. In preliminary experiments, we analyzed ETC protein expression using real-time qPCR. Our data show that treatment with flutamide significantly upregulates the expression of genes involved in mitochondrial ETC, such as mtND6, while decreasing the expression of the lipogenic genes Acly and Acc. These additional data are now presented in Supplementary Figure S3B. (lines 223-226)
(9) 245: Aren't the authors showing in Fig. 5 that glut4 expression is reduced in seminal vesicle epithelial cells upon testosterone treatment? How does that fit into the author's hypothesis?
Thank you for pointing this out. We have already responded to a similar comment from Reviewer 3 in a previous revision. Please refer to our response to Reviewer 3 in a previous version.
(10) 285: Based on the author's results OA increases the oocyte cleavage rate but then reduces the rate of blastocyst to cleaved oocyte. Doesn't that mean OA affects negatively early development?
We thank the reviewer for the insightful comment. The one-hour pre-treatment is designed to reflect the transient exposure of sperm to the seminal plasma during ejaculation. In this context, it is unlikely that such a short exposure would impair the overall developmental potential of the embryo. However, although pre-conditioning with oleic acid does not ultimately affect the development of the offspring, it may lead to a decrease in the blastocyst rate at a certain point (approximately 96-120 hours after fertilization). We agree that additional research is needed to demonstrate this.
Therefore, because the experiments related to the effects of oleic acid on sperm and fertilization are currently incomplete, we have decided to withdraw them for future research.
(11) 305: What happens to pyruvate and lactate levels when ACLY expression is reduced?
We appreciate the reviewer’s question regarding the fate of pyruvate and lactate when ACLY expression is reduced. In the absence of testosterone (Ctrl), the expression level of ACLY decreases. At this time, the concentration of pyruvate in the culture medium increased compared to that of testosterone (Testo; Fig. 4D,E). This is probably a reflection of the fact that when the expression of ACLY is suppressed, the rate at which the products of the glycolytic pathway are converted to the fat-producing pathway (i.e., the conversion of citrate to acetyl-CoA) decreases.
On the other hand, lactate levels did not change significantly. This suggests that the flow of lactate production via lactate dehydrogenase is relatively constant, independent of metabolic reprogramming by ACLY.
Therefore, our data suggest that a decrease in ACLY expression leads to a decrease in pyruvate demand, while lactate production is maintained. We interpret these findings as supporting the idea that ACLY is important for directing the carbon produced by the glycolytic pathway to lipid synthesis (by transporting citrate from the mitochondria).
We hope that this explanation clarifies the interpretation of the data.
Minor revision:
189: ECAR: extracellular acidification rate. Please correct.
We have corrected this. (Lines 184-185)
199: Pyruvate is not synthesized, it is metabolized from PEP. Please correct.
The following corrections have been made. “pyruvate is metabolized from phosphoenolpyruvic acid through glycolysis”. (Lines 194-195)
In addition, minor revisions were made to improve the clarity of the overall text.
-
-
eLife Assessment
Seminal plasma is a crucial component of semen that can affect sperm capacitation. However, the role of seminal plasma components, including fatty acids, in sperm function and fertility is poorly understood. In this important study, the authors provide a solid evidence of the testosterone-induced metabolic shift in the epithelial cells of seminal vesicle to support an fatty acid synthesis and also describe the potential effect of oleic acid on sperm motility.
-
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors identify oleic acid as a particularly important metabolite, derived from seminal vesicle epithelium, that stimulates linear progressive motility in isolated cauda epidydimal sperm in vitro. The authors provide additional experimental evidence of a testosterone dependent mechanism of oleic acid production by the seminal vesicle epithelium.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as add important …
Reviewer #1 (Public review):
Summary:
In this revised report, Yamanaka and colleagues investigate a proposed mechanism by which testosterone modulates seminal plasma metabolites in mice. The authors identify oleic acid as a particularly important metabolite, derived from seminal vesicle epithelium, that stimulates linear progressive motility in isolated cauda epidydimal sperm in vitro. The authors provide additional experimental evidence of a testosterone dependent mechanism of oleic acid production by the seminal vesicle epithelium.
Strengths:
Often, reported epidydimal sperm from mice have lower percent progressive motility compared with sperm retrieved from the uterus or by comparison with human ejaculated sperm. The findings in this report may improve in vitro conditions to overcome this problem, as well as add important physiological context to the role of reproductive tract glandular secretions in modulating sperm behaviors. The strongest observations are related to the sensitivity of seminal vesicle epithelial cells to testosterone. The revisions include addition of methodological detail, modified language to reflect the nuance of some of the measurements, as well as re-performed experiments with more appropriate control groups. The findings are likely to be of general interest to the field by providing context for follow-on studies regarding the relationship between fatty acid beta oxidation and sperm motility pattern.
Weaknesses:
Support for the proposed mechanism is stronger in this revised report than in the previous report, but there are many challenges in measuring sperm metabolism and its direct relationship with motility patterns. This study is no exception and largely relies on correlations between various experiments in lieu of direct testing. Additionally, the discussion is framed from a human pre-clinical perspective, and it should be noted that the reproductive physiology between mice and humans is very different.
-
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. 18:1 oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility …
Reviewer #2 (Public review):
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels as well as isolated mouse and human seminal vesicle epithelial cells the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces a difference in gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes regulating cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid. The revised version strengthens the role of ACLY as the main regulator of seminal vesicle epithelial cell metabolic programming. 18:1 oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility and in vivo fertilization in the presence of 18:1 oleic acid and the absence of testosterone, however, is small. Additional experiments should be included to further support that oleic acid positively affects sperm function.
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
In this report, the authors investigated the effects of reproductive secretions on sperm function in mice. The authors attempt to weave together an interesting mechanism whereby a testosterone-dependent shift in metabolic flux patterns in the seminal vesicle epithelium supports fatty acid synthesis, which they suggest is an essential component of seminal plasma that modulates sperm function by supporting linear motility patterns.
Strengths:
The topic is interesting and of general interest to the field. The study employs an impressive array of approaches to explore the relationship between mouse endocrine physiology and sperm function mediated by seminal components from various glandular secretions of …
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
In this report, the authors investigated the effects of reproductive secretions on sperm function in mice. The authors attempt to weave together an interesting mechanism whereby a testosterone-dependent shift in metabolic flux patterns in the seminal vesicle epithelium supports fatty acid synthesis, which they suggest is an essential component of seminal plasma that modulates sperm function by supporting linear motility patterns.
Strengths:
The topic is interesting and of general interest to the field. The study employs an impressive array of approaches to explore the relationship between mouse endocrine physiology and sperm function mediated by seminal components from various glandular secretions of the male reproductive tract.
Thank you for your positive evaluation of our study's topic and approach. We are pleased that you found our investigation into the effects of reproductive secretions on sperm function to be of general interest to the field. We appreciate your positive feedback on the diverse methods we employed to explore this complex relationship.
Weaknesses:
Unfortunately, support for the proposed mechanism is not convincingly supported by the data, and the experimental design and methodology need more rigor and details, and the presence of numerous (uncontrolled) confounding variables in almost every experimental group significantly reduce confidence in the overall conclusions of the study.
The methodological detail as described is insufficient to support replication of the work. Many of the statistical analyses are not appropriate for the apparent designs (e.g. t-tests without corrections for multiple comparisons). This is important because the notion that different seminal secretions will affect sperm function would likely have a different conclusion if the correct controls were selected for post hoc comparison. In addition, the HTF condition was not adjusted to match the protein concentrations of the secretion-containing media, likely resulting in viscosity differences as a major confounding factor on sperm motility patterns.
We appreciate you highlighting concerns regarding our weak points and apologize for our unclear description. We revised the manuscript to be as rigorous and detailed as possible. In addition, some experimental designs were changed to simpler direct comparisons, and additional experiments were conducted (New Figure 1A-F, lines 103-113). We have made our explanations more consistent with the provided data, which includes further experimentation with additional controls and larger sample sizes to increase the reliability of the findings.
To address the multiple testing problem, a multiple testing correction was made by making the statistical tests more stringent (Please see Statistical analysis in the Methods section and the Figure legends). Based on different statistical methods, the analysis results did not require significant revisions of the previous conclusions.
Because the experiments on mixing extracts from the seminal vesicles were exploratory, we planned to avoid correcting for multiple comparisons. Repeating the t-test could lead to a Type I error in some results, so we apologize for not interpreting and annotating them. In the revised version, we removed the dataset for experiments on mixing extracts from the seminal vesicles and prostate, and we changed the description to refer to the clearer dataset mentioned above.
The viscosity of the secretion-containing medium was measured with a viscometer, confirming that secretions did not significantly affect the viscosity of the solution. In addition, as the reviewer pointed out, we addressed the issue that the HTF condition could not be used as a control because of the heterogeneity in protein concentration (New Fig.1G, lines 110-111).
Overall, we concluded that seminal vesicle secretion improves the linear motility of sperm more than prostate secretion.
There is ambiguity in many of the measurements due to the lack of normalization (e.g. all Seahorse Analyzer measurements are unnormalized, making cell mass and uniformity a major confounder in these measurements). This would be less of a concern if basal respiration rates were consistently similar across conditions and there were sufficient independent samples, but this was not the case in most of the experiments.
We apologize for the many ambiguities in the first manuscript. Cell culture experiments in the paper, including the flux analysis, were performed under conditions normalized or fixed by the number of viable cells. The description has also been revised to emphasize that the measurement values are standardized by cell count (lines 183-185, 189-190, 194-197). We emphasize that testosterone affects metabolism under the same number of viable cells (New Fig.4). This change in basal respiration is thought to be due to the shift in the metabolic pathway of seminal vesicle epithelial cells to a “non-normal TCA cycle” in which testosterone suppresses mitochondrial oxygen consumption, even under aerobic conditions (New Figs.3, 4, 5).
The observation that oleic acid is physiologically relevant to sperm function is not strongly supported. The cellular uptake of 10-100uM labeled oleic acid is presumably due to the detergent effects of the oleic acid, and the authors only show functional data for nM concentrations of exogenous oleic acid. In addition, the effect sizes in the supporting data were not large enough to provide a high degree of confidence given the small sample sizes and ambiguity of the design regarding the number of biological and technical replicates in the extracellular flux analysis experiments.
Thank you for your important critique. As you noted, the too-high oleic acid concentration did not reflect physiological conditions. Therefore, we changed the experimental design of an oleic acid uptake study and started again. We added an in vitro fertilization experiment corresponding to the functional data of exogenous oleic acid at nM concentrations (New Fig.7J,K, Lines 274-282).
For the flux data to determine the effect of oleic acid on sperm metabolism, we have indicated in the text that the data were obtained based on eight male mice and two technical replicates. Pooled sperm isolated and cultured from multiple mice were placed in one well. The measurements were taken in three different wells, and each experiment was repeated four times. We did not use the extracellular flux analyzers XFe24 or XFe96. The measurements were also repeated because the XF HS Mini was used in an 8-well plate (only a maximum of 6 samples at a run since 2 wells were used for calibration).
Overall, the most confident conclusion of the study was that testosterone affects the distribution of metabolic fluxes in a cultured human seminal vesicle epithelial cell line, although the physiological relevance of this observation is not clear.
We thank the comments that this finding is one of the more robust conclusions of our study. Below we have written our thoughts on the physiological relevance of the observation results and our proposed revisions. In the mouse experiments, when the action of androgens was inhibited by flutamide, oleic acid was no longer synthesized in the seminal vesicles. The results of the experiments using cultured seminal vesicle epithelial cells showed that oleic acid was not being synthesized because of a change in metabolism dependent on testosterone. We have also added IVF data on the effects of oleic acid on sperm function (New Fig.7 and Supplementary Fig. 5, lines 274-282).
As you can see, we have obtained consistent data in vitro and in vivo in mice. Our data also showed that the effects of testosterone on metabolic fluxes in vitro are similar in mouse and human seminal vesicle epithelial cells (New Fig.9). Therefore, it can be assumed that a decrease in testosterone levels causes abnormalities in the components of human semen. However, the conclusion was overestimated in the original manuscript, so we changed the wording as follows: It could be assumed that a decrease in testosterone levels causes abnormalities in the components of human semen. (lines 422-423)In the introduction, the authors suggest that their analyses "reveal the pathways by which seminal vesicles synthesize seminal plasma, ensure sperm fertility, and provide new therapeutic and preventive strategies for male infertility." These conclusions need stronger or more complete data to support them.
We appreciate your comments about the suggestion presented in the introduction.
We also removed our conclusions regarding treatment and prevention strategies for male infertility (lines 96-98). We wanted to discuss our findings not conclusively but as future applications that could result from further research based on our initial findings.
The last sentence of the introduction has been revised to tone down these assertions as follows: These analyses revealed that testosterone promotes the synthesis of oleic acid in seminal vesicle epithelial cells and its secretion into seminal plasma, and the oleic acid ensures the linear motility and fertilization ability of sperm.
We are grateful for your suggestions, which have prompted us to refine our manuscript.
Reviewer #2 (Public Review):
Summary:
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels, as well as isolated mouse and human seminal vesicle epithelial cells, the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces differential gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes that regulate cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid.
Strength:
Oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility and in vivo fertilization in the presence of 18:1 oleic acid and the absence of testosterone is small but significant, suggesting that the authors have identified one of the fertilization-supporting factors in seminal plasma.
Thank you for your positive comments regarding our work on the role of testosterone in regulating metabolic enzymes and the subsequent production of 18:1 oleic acid in seminal vesicle epithelial cells. We are pleased that the strength of our findings, particularly identifying oleic acid as a factor influencing sperm motility and mitochondrial respiration, has been recognized.
Weaknesses:
Further studies are required to investigate the effect of other seminal vesicle components on sperm capacitation to support the author's conclusions. The author's experiments focused on potential testosterone-induced changes in the rate of seminal vesicle epithelial cell glycolysis and oxphos, however, provide conflicting results and a potential correlation with seminal vesicle epithelial cell proliferation should be confirmed by additional experiments.
Thank you very much for your valuable criticism. Although we fully agree with your comment, conducting experiments to investigate the effects of other seminal vesicle components on the fertilization potential of sperm would be a great challenge for us. This is because it has taken us the last three years to identify oleic acid as a key factor in seminal plasma. We are considering a follow-up study to explore the effect of other seminal vesicle components on sperm capacitation. Therefore, we have revised the Introduction and conclusions to tone down our assertions .
The revised manuscript also includes additional data showing a correlation between changes in metabolic flux and the proliferation of seminal vesicle epithelial cells using shRNA. As a result, it was shown that cell proliferation is promoted when mitochondrial oxidative phosphorylation is promoted by ACLY knockdown (New Fig.8D, lines 303-305). This shows a close relationship between the metabolic shift in seminal vesicle epithelial cells and cell proliferation. The revised manuscript includes an interpretation and discussion of these results (lines 369-379).
We are grateful for your suggestions, which have prompted us to refine our manuscript.
Reviewer #3 (Public Review):
Summary:
Male fertility depends on both sperm and seminal plasma, but the functional effect of seminal plasma on sperm has been relatively understudied. The authors investigate the testosterone-dependent synthesis of seminal plasma and identify oleic acid as a key factor in enhancing sperm fertility.
Strengths:
The evidence for changes in cell proliferation and metabolism of seminal vesicle epithelial cells and the identification of oleic acid as a key factor in seminal plasma is solid.
Weaknesses:
The evidence that oleic acids enhance sperm fertility in vivo needs more experimental support, as the main phenotypic effect in vitro provided by the authors remains simply as an increase in the linearity of sperm motility, which does not necessarily correlate with enhanced sperm fertility.
We appreciate the positive feedback on the solid evidence of cell proliferation and metabolic changes in seminal vesicle epithelial cells and the identification of oleic acid as an important factor in seminal plasma. We fully agree with the assessment that the evidence linking oleic acid and increased sperm fertility in vivo needs further experimental support. To address this concern, we changed the experimental design of an oleic acid study and started again to be more physiological regarding the effect of oleic acid on fertility outcomes, increased the replicates of artificial insemination, and added in vitro fertilization assessments (New Fig.7 and supplementary Fig.5, lines 274-282). The revised manuscript describes these experiments and discusses the association between oleic acid and fertility.
We are grateful for your suggestions, which have prompted us to refine our manuscript.
Recommendations for the authors:
Reviewing Editor's note:
As you can see from the three reviewers' comments, the reviewers agree that this study can be potentially important if major concerns are adequately addressed. The major concern common to all the reviewers is the incomplete mechanistic link between the physiological androgen effect on the production of oleic acid and its effect on sperm function. Statistical analyses need more rigor and consideration of other important capacitation parameters are needed to address these concerns and to improve the manuscript to support the current conclusions.
Thank you for summarizing the reviewers' feedback and for your insights regarding the major concerns raised. We appreciate the reviewers' understanding of the potential importance of our work and have addressed the issues highlighted to strengthen the manuscript. We believe these changes will improve the quality of the manuscript and provide a clearer and more complete understanding of the role of androgens and oleic acid in sperm function.
Reviewer #1 (Recommendations For The Authors):
The following comments are provided with the hope of aiding the authors in improving the alignment between the data and their interpretations.
Thank you for allowing us to strengthen our manuscript with your valuable comments and queries. We have made our best efforts to reflect your feedback.
Major Comments:
(1) The methodological detail is not sufficient to reproduce the work. For example:
a. Manufacturer protocols are referred to extensively. These protocols are neither curated nor version-controlled. Please consider describing the underlying components of the assays. If information is not available, please consider providing catalog numbers and lot numbers in the methods (if appropriate for journal style requirements).We appreciate this suggestion, which we believe is important to ensure reproducibility. We described the catalog number in our Methodology and included as much information as possible.
b. Please consider describing the analyses in full, with consideration given to whether blinding was part of the design. For example- line 492: "apoptotic cells were quantified using ImageJ". How was this quantified? How were images pre-processed? Etc.
Although blinding was not performed, experiments and analyses based on Fisher's three principles were conducted to eliminate bias (lines 549-552). In order to avoid false-positive or false-negative results, it is clearly stated that tissue sections treated with DNAse were used as positive controls, and tissue sections without TdT were used as negative controls for apoptosis. We have added detailed quantification methods (lines 544-546).
c. Please consider providing versions of all acquisition and analysis software used.
We have added software version information in Materials and Methods.
(2) Please consider revisiting the statistical analyses. Many of the analyses don't seem appropriate for the design. For example, the use of a t-test with multiple comparisons for repeated measures design in Figure 2 and the use of t-test for two-factor design in Figure 8. etc.
To address the multiple testing issues, the statistical methodology was changed to a more rigorous one. Details are given in the Statistical analysis in the Methods section and the Figure legends.
(3) The increase in % LIN in Figure 1 may be confounded by differences in viscosity between HTF and the fluid secretion mixtures. For this reason, HTF may not be an appropriate control for the ANOVA post hoc analysis. HTF protein was not adjusted to the same concentration as the secretion mixtures, correct? Ultimately, it does not appear that there would be a significant statistical effect of the different fluid mixtures if appropriate statistical comparisons were made. This detracts from the notion that the secretions impact sperm function.
(4) Figure 1, the statistical analysis in the legend suggests that the experiments were analyzed with a t-test. Were corrections made for multiple comparisons in B-D? An ANOVA would probably be more appropriate.
We used a viscometer to measure the viscosity of a solution of prostate and seminal vesicle secretions adjusted to a protein concentration of 10 mg/mL. The results showed that the secretions did not cause any significant viscosity changes (New Fig.1G, Lines 110-111).
As you pointed out, the protein levels in the HTF medium and the secretion mixture are not adjusted to the same concentration. In addition, the original manuscript was not a controlled experiment because the two factors, seminal vesicle and prostate extracts, were modified. Therefore, to investigate the effect of prostate and seminal vesicle secretions on sperm motility, we modified the experimental design to directly compare the effects of the two groups: seminal vesicle and prostate extracts (New Fig.1A-G, lines 101-113). To show the sperm quality used in this study, motility data from sperm cultured in the HTF medium are presented independently in New Supplemental Fig.1A.
(5) Additionally in Figure 1, there is no baseline quality control data to show that there are no intrinsic differences between sperm sampled from the two treatment groups. So baseline differences in sperm quality/viability remain a potential confounder.
We thank you for this important point. Epididymal sperm were collected from healthy mice. We recovered only the seminal vesicle secretions from the flutamide-treated mice to pursue its role in the accessory reproductive glands, since testosterone targets the testes and accessory reproductive organs. So, there was no qualitative difference between the epididymal sperm before treatment. Nevertheless, incubation with seminal vesicle secretion for one hour altered the sperm motility pattern and in vivo fertilization results. Sperm function was altered by seminal vesicle secretion in a short period of culture time. We apologize for the confusion, and we have revised the text and figure to carry a clearer message (lines 128-132).
(6) Figure 1E, did the authors confirm that flutamide-treated mice had decreased serum androgens? How often were mice treated with flutamide? This is important because flutamide has a relatively short half-life and is rapidly metabolized to inert hydroxyflutamide.
Serum testosterone levels were unchanged. Flutamide was administered every 24 hours for 7 consecutive days. Although there was no change in blood testosterone levels (New Supplemental Fig.1B), a decrease in the weight of the seminal vesicles, prostate, and epididymis was confirmed. This is thought to be due to the pharmacological activity of flutamide.
(7) Figure 1H, the meaning of 'relative activity of mitochondria' isn't clear. JC-1 does not measure 'activity'. A decreased average voltage potential across the inner mitochondrial membrane may indicate that more of the sperm from the flutamide group were dead. Additionally, J-aggregates are slow to form, generally requiring long incubation periods of at least 90 minutes or more. Additional positive and negative controls for predictable mitochondrial transmembrane voltage potential polarization states would have improved the quality of this experiment.
Thank you for pointing this out. We have replaced the relative activity of mitochondria with high mitochondrial membrane potential (New Fig.1M, lines 125-128). Actually, it is thought that the sperm cultured in seminal vesicle secretions from mice that had been administered flutamide died because the motility of the sperm was also significantly reduced. Since antimycin reduces mitochondrial membrane potential, we have added an experiment in which 10 µM antimycin-treated sperm were used as a control to confirm that the JC-1 reaction is sensitive to changes in membrane potential.
(8) Figure 4, the extracellular flux data appear to be unnormalized. The Seahorse instruments are extremely sensitive to the mass and uniformity of the cells at the bottom of the well. This may be a significant confounder in these results. For example, all of the observed differences between groups could simply be a product of differential cell mass, which is in line with the reduced growth potential of testosterone-treated cells indicated by the authors in the results section.
We thank you for this important point. After correcting for cell viability, we seeded the same number of viable cultured cells into wells between experimental groups before measuring them in the flux analyzer. There were no significant differences in survival rates in all experiments. As a result, an increase in glucose-induced ECAR and a suppression of mitochondrial respiration were observed. We would like to emphasize that this difference based on metabolic data does not imply a reduction in the growth potential of the cells due to testosterone treatment.
We described that these measurements are normalized based on cell count and viability (lines 184, 190, 195).
(9) How did the authors know that the isolated mouse primary cells were epithelial cells? Was this confirmed? What was the relative sample purity?
The cells were labeled with multiple epithelial cell markers (cytokeratin) and confirmed using immunostaining and flow cytometry. The percentage of cells positive for epithelial cell markers was approximately 80%. A stromal cell marker (vimentin) was also used to confirm purity, but only a few percent of cells were positive. The contaminating cell type was considered to be mainly muscle cells because the gene expression levels of muscle cell markers verified by RNA-seq were relatively high.
(10) It is misleading to include the lactate/pyruvate media measurements in the middle of the figure in Figure 4 D and E because it seems at first glance like these measurements were made in the seahorse media but they are completely unrelated. Additionally, these measures are not normalized and are sensitive to confounding differences in cell viability, seeding density, mass, etc.
Thank you for pointing this out. We have placed the lactate and pyruvate measurement graphs after the flux data of ECAR. We noted that these measurements are normalized based on cell count and viability (lines 189-190). The doubling time of seminal vesicle epithelial cells was approximately 3 days, and testosterone inhibited cell proliferation. Therefore, the seeding concentration of cells was increased 4-fold in the testosterone-treated group compared to the control, and experiments were conducted to ensure that the confluency at the time of measurement after 7 days of culture was comparable between groups.
(11) The flux analyzer assays sold by Agilent have many ambiguities and problems of interpretation. Unfortunately, Agilent's interest in marketing/sales has outpaced their interest in scientific rigor. Please consider revising some of the language regarding the measurements. For example, 'ATP production rate' is not directly measured. Rather, oligomycin-sensitive respiration rate is measured. The conversion of OCR to ATP production rate is an estimation that depends on complex assumptions often requiring additional testing and validation. The same is true for other ambiguous terms such as 'maximal respiration' referring to FCCP uncoupled respiration, and glycolytic rate- which is also not measured directly. If the authors are interested in a more detailed description of the problems with Agilent's interpretation of these assays please see the following reference (PMID: 34461088).
Thank you for your critical criticism and thoughtful advice, as well as for sharing the excellent reference. We agree with you on the flux analyzer ambiguities and data interpretation problems. The description of the measured values has been revised as follows.
We have replaced the “ATP production rate” with the “oligomycin-sensitive respiratory rate.” Similarly, we have replaced “maximal respiration” with “FCCP-induced unbound respiration.” (lines 197-202) We chose not to deal with the conversion of OCR to ATP production rate because it is outside the scope of interest in our study.
Avoid using the term "glycolytic capacity". We use “Oligomycin-sensitive ECAR.” (line 186) We recognize that the ECARs measured in this study reflect experimental conditions and may not fully represent physiological glycolytic flux in vivo. So, the main section includes a data set of glucose uptake studies to emphasize the significance of the changes obtained with the flux analyzer assay. (New Fig.6, lines 230-254)
Figure 6, it's not surprising to see the accumulation of labeled oleic acid in the cells, however, this does not mean that oleic acid is participating in normal metabolic processes. Oleic acid will have detergent effects at high (uM) concentrations. The observation that sperm 'take up' OA at 10-100 uM concentrations should also be validated against sperm function the health of the cells is very likely to be negatively impacted. Additionally, no apparent accumulation is noted in the fluorescence imaging at 1uM, but the authors insinuate that uptake occurs at low nM concentrations. The effects in Figure 6D-F are nominal at best and are likely a result of the small sample sizes.
Thank you for your good suggestion. We agree with the reviewer that high concentrations of oleic acid had a detergent effect. To improve the consistency of functional data and observations, oleic acid uptake tests were performed under the same concentration range as the sperm motility tests (New Fig.7A-C). The oleic acid concentration at this time was calculated regarding the oleic acid concentration in seminal fluid recovered from mice as detected by GCMS to reflect in vivo conditions.
Epididymal sperm were incubated with fluorescently labeled oleic acid and observed after quenching of extracellular fluorescence. Fluorescent signals were detected selectively in the midpiece of the sperm. The fluorescence intensity of sperm quantified by flow cytometry increased significantly in a dose-dependent manner (New Fig.7A-C, lines 261-264).
Furthermore, increasing the sample size did not change the trend of the sperm motility data. Although the effect size of oleic acid on sperm motility was small (New Fig.7D-G, lines 265-268), an improvement in fertilization ability was observed both in vitro (IVF) and in vivo (AI) (New Fig.7J-L, lines 274-282, 286-291). We conclude that the effect of oleic acid on sperm is of substantial significance. These data and interpretations have been revised in the text in the Results section.
(12) Figure 6H, I applaud the authors for attempting intrauterine insemination experiments to test their previous findings. That said, there is no supporting data included to show that the sperm from the treatment groups had comparable starting viability/quality. Additionally, it is difficult to tell if the results are due to the small sample sizes and particularly the apparent outlier in the flutamide-only group.
Thanks for the praise and comments for improvement. As we answered in your comment #5 above, the epididymal sperm was collected from healthy mice. Therefore, there is no qualitative difference in the epididymal sperm before treatment. This is described in the figure legend (lines 1130-1131). We apologize again for this complication. We also more than doubled the number of replications of the experiment. The impact of the outlier would have been minimal.
(13) One final question related to Figure 6H: how did the authors know they were retrieving all of the possible 2-cell embryos from the uterus? Perhaps the authors could provide the raw counts of unfertilized eggs and 2-cell embryos so we can see if there were differences between the mice.
We retrieved the pronuclear stage embryos from the fallopian tubes. It is not certain whether all embryos were recovered. Therefore, we added the number of embryos in the graph and in the supplementary data.
(14) Figure 7 has the same seahorse assay normalization problem as mentioned earlier. Without normalization, it is difficult to tell if the effects are simply due to differences in cell mass. Were the replicates indicated in the graphs run on the same plate? If so, it would be much more convincing to see a nested design, with technical replicates within plates, and additional replicates run on separate plates.
As we answered in your comment #8 above, these measurements were normalized based on sperm count. This has been corrected to be noted in the text and the figure legend (lines 1123-1124).
Pooled sperm isolated and cultured from multiple mice were placed in one well. The measurements were taken in three different wells, and each experiment was repeated four times. We did not use the extracellular flux analyzers XFe24 or XFe96. The measurements were also repeated because the XF HS Mini was used in an 8-well plate (only a maximum of 6 samples at a run since 2 wells were used for calibration).
(15) The statistical test in Figures 8E and F described in the legend is inappropriate (t-test), this appears to be a two-factor design.
Thank you for pointing this out. Differences between groups were assessed using a two-way analysis of variance (ANOVA). When the two-way ANOVA was significant, differences among values were analyzed using Tukey's honest significant difference test for multiple comparisons.
(16) The data in Figure 8 are interesting, and the effects appear to be a little more consistent compared with the mouse primary cells, potentially due to cell uniformity. However, the data are unnormalized, causing significant ambiguity, and there are no measures of cell viability to determine if the effects are due to cell death (or at least relative cell mass).
As we answered in your comments #8 and #14 above, these measurements were normalized based on cell count and viability. This has been corrected to be noted in the figure legend (lines 1185-1186).
Minor Comments:
(1) The section title indicating the beginning of the results section is missing.
A section title has been added to indicate the beginning of the results section.
(2) There were several typos and confusingly worded statements throughout. Please consider additional editing.
We used a proofreading service and corrected as much as possible.
(3) In the introduction, a brief description of seminal fluid physiology is provided, but the reference is directed toward human physiology. Given that the research is performed solely in the mouse, a brief comparative description of mouse physiology would be helpful. For example, what is the role of mouse seminal fluid in the formation of the mating plug? What are the implications of the relative size disparity in seminal vesicles in mice versus humans? Etc.
The third paragraph of the introduction has been revised (lines 57-60).
Reviewer #2 (Recommendations For The Authors):
Thank you for allowing us to strengthen our manuscript with your valuable comments and queries. We have made our best efforts to reflect your feedback.
(1) The abstract is confusing and partly misleading and should be revised to more clearly and accurately summarize the study.
The abstract was revised to be clearer and more accurate (lines 20-34).
(2) The introduction should be revised to more accurately describe the sperm life cycle. Spermatogenesis, per definition, for example, exclusively takes place in the testis, sperm do not gain fertilization competence in the epididymis, sperm isolated from the epididymis cannot fertilize an oocyte unless in vitro capacitated, etc. In the last paragraph the connection between changes in fructose and citrate concentration, sperm metabolism and testicular-derived testosterone and AR remain unclear.
The introduction was revised to be clearer and more accurate (lines 44-45).
Citric acid and fructose are chemical components that are the subject of biochemical testing and are commonly used as semen testing items for humans and livestock. This is because the secretory function of the prostate and seminal vesicles is dependent on androgens. The measurement of citric acid and fructose concentrations in semen is routinely used to indicate testicular androgen production function (ISBN: 978-1-4471-1300-3, 978 92 4 0030787).
(3) Throughout the manuscript the concept of (in vitro) capacitation is missing. Mixing sperm with seminal plasma is not the only way to achieve sperm that can fertilize the oocyte. Since media containing bicarbonate and albumin is the standard procedure in the field to capacitate epididymal mouse sperm rein vitro, the manuscript would gain value from a comparison between the effect of seminal plasma and in vitro capacitating media. Interesting readouts in addition to motility would i.e. be sAC activation, PKA-substrate phosphorylation, and acrosomal exocytosis.
Thank you for pointing out this important point. As the reviewer points out, fertilization can be achieved in artificial insemination and in vitro fertilization using epididymal sperm which have not been exposed to seminal plasma. This has historically led to an underestimation of the role of accessory reproductive glands, such as the prostate and seminal vesicles. However, it has been reported that the removal of seminal vesicles in rodents decreases the fertilization rate after natural mating. This has been shown to be due to multiple factors affecting sperm motility rather than factors involved in plug formation (PMID: 3397934), but details of these factors and the whole picture of the role of the accessory glands were not known. This led us to become interested in the effects of sperm plasma on sperm other than fertilization and led us to begin research on the role of the accessory glands that synthesize sperm plasma.
Early in our study, we found that simply exposing sperm to seminal vesicle extracts for 1 hour before IVF dramatically reduced fertilization rates, even in HTF medium containing bicarbonate and albumin. The experiment was designed on the assumption that seminal plasma contains factors that inhibit sperm from acquiring fertilizing ability. Therefore, we conducted experiments using modified HTF without albumin to avoid unintended motility patterns.
However, we also respect the reviewer's opinion, and we have added our preliminary data related to IVF (New supplementary Fig.5).
(4) In the introduction and throughout the manuscript it is unclear what the authors mean by "linear motility". An increase in VSL doesn't mean that the sperm swim in a more linear or straight way, or even that the sperm are 'straightened', it means that they swim faster from point A to point B. Do the authors mean progressive or hyperactivated motility? Please clarify.
For all conditions tested the authors should follow the standard in the field and include the % of motile, progressively motile, and hyperactivated sperm.
Thank you for pointing this out. We appreciate your feedback regarding the terminology. In our manuscript, "linear motility" refers to the degree to which sperm move in a straight line. We have clarified this by explaining that VSL (Straight-Line Velocity) and LIN (Linearity) are used to quantify and describe linear motility in sperm analysis: Higher VSL values indicate more direct, linear movement. A higher LIN value indicates a straighter path, thus representing greater linear motility. These terms have been standardized, and explanations have been added to the main text (lines 111-113).
In response to your suggestion, we have included the percentage of motility and progressive motility for all conditions tested. However, since the experiment was performed using modified HTF without albumin, we have decided not to report the percentage of hyperactivation to avoid confusion.
(5) Did the authors confirm that the injection of flutamide decreases androgen levels? That control needs to be included in the experiment to validate the conclusion.
Injection of flutamide did not reduce androgen levels (see reviewer #1, comment 6). This is because flutamide's mechanism of action is based on antagonizing androgen and inhibiting its binding to the androgen receptor (New Fig.2A).
(6) The role of mitochondrial activity in sperm progressive motility is still under investigation. PMID: 37440924 i.e. showed that inhibition of the ETC does not affect progressive but hyperactivated motility. The authors should either include additional experiments to confirm the correlation between mitochondrial activity and sperm progressive motility or tone down that conclusion.
We have previously shown that treatment with D-chloramphenicol, an inhibitor of mitochondrial translation, significantly reduced sperm mitochondrial membrane potential, ATP levels, and linear motility (PMID: 31212063). Also, in the previous manuscript, we did not address progressive motility or hyperactivated motility in our analysis. We have chosen to discuss the effect of mitochondrial activity on linear motility rather than on progressive motility and hyperactivation of sperm.
Was mitochondrial activity also altered in epididymal sperm incubated with and without seminal plasma or in aged mice?
The mitochondrial membrane potential of epididymal sperm cultured with seminal vesicle extract (SV) was higher than that of epididymal sperm cultured without seminal vesicle extract (without SV: 67.3 ± 0.8%, with SV: 83.4 ± 1.8%). On the other hand, the mitochondrial membrane potential of epididymal sperm cultured with seminal vesicle extract recovered from aged mice was decreased (SV from aged: 60.3 ± 2.7%). It should be noted that the epididymal spermatozoa used in these experiments were healthy individuals, different from those from which seminal vesicle extracts were collected. (See also the response to reviewer 1's comment #5.)
(7) The quality of the provided images showing AR, Ki67, and TUNEL staining should be improved or additional images should be included. Especially the AR staining is hard to detect in the provided images. The authors should also include a co-staining between AR and vesicle epithelial cells. That epithelial cells are multilayered does not come across in the pictures provided.
We apologize for any inconvenience caused. The image has been replaced with one of higher resolution. The multilayered structure of the epithelial cells will also be seen.
For the 12-month-old mice, an age-matched control should be included to support the authors' conclusion.
To clarify the seminal vesicle changes associated with aging, we included images of 3-month-old mice as controls (New Supplementary Fig.2D).
Overall, the rationale for the experiment does not become clear. How are the amount of seminal vesicle epithelial cells, testosterone, and AR expression connected to seminal plasma secretions? Why is it a disadvantage to have proliferating seminal vesicle epithelial cells? How is proliferation connected to the proposed switch in metabolic pathway activity?
We have added some explanations and supporting data to the manuscript (New Fig.8D, lines 303-305, 315-319, 369-379). Cell proliferation stopped when the metabolic shift occurred, redirecting glucose toward fatty acid synthesis. Fatty acid synthesis is an important function of the seminal vesicle, and in the presence of testosterone, fatty acid synthesis enhancement and arrest of proliferation occur simultaneously. The connection between metabolism and cell proliferation was further demonstrated when ACLY was knocked down by shRNA, which stopped fatty acid synthesis and released the proliferative arrest induced by testosterone, allowing the cells to proliferate again. However, we do not know what effects occur when cell proliferation is stopped.
(8) The experiments provided for glycolysis and oxphos are inconsistent and insufficient to support the authors' conclusion that testosterone shifts glycolytic and oxphos activity of seminal vesicle epithelial cells. Multiple groups (PMID 37440924, 37655160, 32823893) have shown that the increased flux through central carbon metabolism during capacitation is accompanied by an accumulation of intracellular lactate and increased secretion of lactate into the surrounding media. How do the authors explain that they see an increase in glucose uptake and ECAR but not in lactate and a decrease in pyruvate? Did the authors additionally quantify intracellular pyruvate and lactate? Since pyruvate and lactate are in constant equilibrium, it is odd that one metabolite is changing and the other one is not.
Thank you for pointing this out. Since ECAR is often used as an alternative to lactate production but does not directly measure lactate levels, we measured changes in lactate and pyruvate concentrations in the culture medium. Under our experimental conditions, glucose appeared to be directed primarily towards anabolic processes, such as fatty acid synthesis, rather than the OXPHOS pathway, which may explain the lack of lactate production. The observed decrease in pyruvate might indicate its conversion to acetyl-CoA in the mitochondria, supporting both fatty acid synthesis and the TCA cycle. This shift would be consistent with the metabolic reprogramming toward anabolic activity.
What do the authors mean by "the glycolytic pathway was not enhanced despite the activation of glycolysis" Seahorse, especially using a series of pathway inhibitors, only provides an indirect measurement of glycolysis and oxphos since the instrument does not provide a distinction from which pathways the detected protons are originating. The authors should consider a more optimized experimental design, i.e. the authors could monitor ECAR and OCR in the presence of glucose over time with and without the addition of testosterone. That would be less invasive since the sperm are not starved at the beginning of the experiment and would provide a more direct read-out. Did the authors normalize cell numbers in their experiment? Alternatively, the authors could consider performing metabolomics experiments.
I agree with the reviewer. Buzzwords such as “glycolytic capacity” simply do not make sense, so we have removed them from the phrases noted by the reviewer. Please refer to the response to some of reviewer 1's points regarding the ambiguity of the data measured by the flux analyzer. Nevertheless, the assay design of the flux analysis could be used as a good “starting point” and provide information on the glycolytic system and respiratory control. Therefore, the interpretation of the flux analysis is supported by subsequent data sets.
(9) The authors would strengthen their results by confirming their gene expression data by quantifying the expression of the respective proteins.
Does testosterone treatment increase GLUT4 protein levels in isolated seminal vesicle epithelial cells? Or does it change the localization of the transporter? Are GLUT4 gene and protein levels altered in flutamide-treated cells? How do the authors explain that testosterone increases glucose uptake without changing Glut gene expression?
We performed Western blot analysis to measure GLUT4 protein levels in seminal vesicle epithelial cells after testosterone treatment. The results showed that testosterone does not alter the expression of GLUT4 protein but simply changes its subcellular localization (New Fig.6C,D, lines 238-244).
The discussion includes the interpretation of the observation that testosterone increases glucose uptake by altering localization without altering GLUT4 gene expression, a phenomenon commonly seen in other cells, such as cardiomyocytes (lines 362-364). The revised main figure also includes a data set of changes in GLUT4 localization, including flutamide-treated data. See also Reviewer 3's main comment #1.
(10) Considering that the authors claim that SV secretions are crucial for sperm fertilization capacity, how do they explain that fertilization rates are still at 40 % when sperm are treated with flutamide?
It is actually about 50% fertilized with HTF because it is fertilized without SV. Considering this baseline, we found that seminal vesicle secretions positively affect sperm in vivo fertilization. On the other hand, seminal plasma from flutamide-treated mice reduced the fertilization ability of healthy sperm. These are described in the text (lines 283-294).
(11) It would be beneficial for the reader to include a schematic summarizing the results.
Thank you for your advice from the reader's point of view. We have visualized the summaries of this study and added them to the manuscript (New Fig.10).
Minor comments:
Line 38: Male fertility, no article, please revise.
I have changed “The male fertility” to “Male fertility” and added some references (lines 42-43).
Line 55: Seminal plasma or TGFb? Please clarify.
Corrected as follows. “TGFβ, a component of seminal plasma, increases antigen-specific Treg cells in the uterus of mice and humans, which induces immune tolerance, resulting in pregnancy.” (lines 60-62)
Line 63: Why do the authors find it surprising that blood and seminal plasma have different compositions?
This is because seminal plasma contains unique biochemical components that are not normally found in blood or only in small quantities. The intention was to emphasize the unique function of seminal plasma in supporting the physiological functions of sperm and to highlight its complex role by comparing it to blood. We clarified these intentions and reflected them in the revised text (lines 62-67).
Line 94: The headline causes confusion. Seminal plasma does not induce sperm motility, it increases progressive sperm motility.
Corrected as follows. “The effect of androgen-dependent changes in mouse seminal vesicle secretions on the linear motility of sperm” (lines 101-102)
Reviewer #3 (Recommendations For The Authors):
Thank you for allowing us to strengthen our manuscript with your valuable comments and queries. We have made our best efforts to reflect your feedback.
Major:
Figure 4 and Figure 5: The trend shows that GLUT3 is up-regulated and GLUT4 is downregulated although both of them are not statistically significant. However, GLUT4 is picked for all the following experiments based on protein localization. Providing other evidence/discussion why not to further consider other GLUTs will help to justify. Also, this reviewer suggests including GLUT4 localization data in the main figure as it is important data for the logical flow to link the following figures.
We focused on GLUT4 because it was known that testosterone increases glucose uptake by changing the localization of GLUT4 without changing its expression (lines 230-231). In the revised manuscript, the increasing trend in Glut3 gene expression was also mentioned in the discussion, in addition to GLUT4 (lines 360-362). In any case, the results showed that testosterone increased glucose uptake by regulating the function of glucose transporters.
Immunostaining of GLUT1~4 was performed to compare seminal vesicles from flutamide-treated mice with controls, and localization changes were observed only in GLUT4. Therefore, we hypothesized that GLUT4 is regulated by testosterone and performed the experiment. Fortunately, we were able to obtain a GLUT4-specific inhibitor, which dramatically inhibited the testosterone-dependent glucose uptake and subsequent lipid synthesis in seminal epithelial cells, leading us to believe that GLUT4 is a major glucose transporter.
Increasing sperm linearity by oleic acid is observed and interpreted as enhanced sperm fertilizing potential. It is not clear why and how sperm linearity can be a determinant factor for enhancing sperm fertility in vivo. Providing an explanation of the effect of oleic acid on another key motility parameter more proven to be directly correlated with fertility (i.e., hyperactivation), and more direct evidence of oleic acid on enhancing sperm linearity indeed increasing sperm fertilization using IVF, is strongly recommended to support the author's main conclusion.
Thank you for pointing this out. It is known that proteins derived from the seminal vesicles inhibit the hyperactivation of sperm and the acrosome reaction. Therefore, we conducted an experiment to add oleic acid, focusing on fatty acid synthesis caused by the metabolic shift of the seminal vesicles, which had not been known until now.
Sperm were pretreated with an oleic acid-containing medium before IVF and oleic acid enhanced sperm linearity. When the sperm number was sufficient, there was no change in the cleavage rate after in vitro fertilization, but when the sperm count was reduced to one-tenth of the normal, the cleavage rate increased compared to the control (lines 274-282). In other words, the physiological role of oleic acid is to increase the probability of fertilization by keeping the sperm motility pattern linear or progressive. This increases the likelihood of the sperm passing through the female reproductive tract and environments that are unfavorable to sperm survival. Our research has uncovered significant insights into the role of seminal vesicle fluid and oleic acid in sperm fertilization. Due to the strong effect of the decapacitation factor, we found that seminal vesicle fluid reduces the fertilization rate in IVF. However, it does not interfere with the fertilization rate in in vivo during artificial insemination. This emphasizes the importance of oleic acid, along with other protein components of seminal plasma, in ensuring the in vivo fertilization ability of sperm.
Minor:
Please correct a typo in Line 173: sifts to shifts
All typographical errors have been corrected.
-
-
eLife assessment
This study reports a potentially important discovery that testosterone-induced metabolic changes in seminal vesicle epithelial cells lead to the production of oleic acids in seminal plasma to enhance sperm fertility. The evidence to support metabolic changes in seminal vesicles and the identification of oleic acid as a key factor in seminal plasma is solid. However, the evidence for how oleic acids support enhanced sperm fertility in vivo is not well supported, thus currently remains incomplete, and requires further study.
-
Reviewer #1 (Public Review):
Summary:
In this report, the authors investigated the effects of reproductive secretions on sperm function in mice. The authors attempt to weave together an interesting mechanism whereby a testosterone-dependent shift in metabolic flux patterns in the seminal vesicle epithelium supports fatty acid synthesis, which they suggest is an essential component of seminal plasma that modulates sperm function by supporting linear motility patterns.
Strengths:
The topic is interesting and of general interest to the field. The study employs an impressive array of approaches to explore the relationship between mouse endocrine physiology and sperm function mediated by seminal components from various glandular secretions of the male reproductive tract.
Weaknesses:
Unfortunately, support for the proposed mechanism is not …
Reviewer #1 (Public Review):
Summary:
In this report, the authors investigated the effects of reproductive secretions on sperm function in mice. The authors attempt to weave together an interesting mechanism whereby a testosterone-dependent shift in metabolic flux patterns in the seminal vesicle epithelium supports fatty acid synthesis, which they suggest is an essential component of seminal plasma that modulates sperm function by supporting linear motility patterns.
Strengths:
The topic is interesting and of general interest to the field. The study employs an impressive array of approaches to explore the relationship between mouse endocrine physiology and sperm function mediated by seminal components from various glandular secretions of the male reproductive tract.
Weaknesses:
Unfortunately, support for the proposed mechanism is not convincingly supported by the data, and the experimental design and methodology need more rigor and details, and the presence of numerous (uncontrolled) confounding variables in almost every experimental group significantly reduce confidence in the overall conclusions of the study.
The methodological detail as described is insufficient to support replication of the work. Many of the statistical analyses are not appropriate for the apparent designs (e.g. t-tests without corrections for multiple comparisons). This is important because the notion that different seminal secretions will affect sperm function would likely have a different conclusion if the correct controls were selected for post hoc comparison. In addition, the HTF condition was not adjusted to match the protein concentrations of the secretion-containing media, likely resulting in viscosity differences as a major confounding factor on sperm motility patterns.
There is ambiguity in many of the measurements due to the lack of normalization (e.g. all Seahorse Analyzer measurements are unnormalized, making cell mass and uniformity a major confounder in these measurements). This would be less of a concern if basal respiration rates were consistently similar across conditions and there were sufficient independent samples, but this was not the case in most of the experiments.
The observation that oleic acid is physiologically relevant to sperm function is not strongly supported. The cellular uptake of 10-100uM labeled oleic acid is presumably due to the detergent effects of the oleic acid, and the authors only show functional data for nM concentrations of exogenous oleic acid. In addition, the effect sizes in the supporting data were not large enough to provide a high degree of confidence given the small sample sizes and ambiguity of the design regarding the number of biological and technical replicates in the extracellular flux analysis experiments.
Overall, the most confident conclusion of the study was that testosterone affects the distribution of metabolic fluxes in a cultured human seminal vesicle epithelial cell line, although the physiological relevance of this observation is not clear.
In the introduction, the authors suggest that their analyses "reveal the pathways by which seminal vesicles synthesize seminal plasma, ensure sperm fertility, and provide new therapeutic and preventive strategies for male infertility." These conclusions need stronger or more complete data to support them.
-
Reviewer #2 (Public Review):
Summary:
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels, as well as isolated mouse and human seminal vesicle epithelial cells, the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces differential gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes that regulate cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid.
Strength:
Oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility and in vivo fertilization in the presence of 18:1 oleic acid and the absence of testosterone is small but …
Reviewer #2 (Public Review):
Summary:
Using a combination of in vivo studies with testosterone-inhibited and aged mice with lower testosterone levels, as well as isolated mouse and human seminal vesicle epithelial cells, the authors show that testosterone induces an increase in glucose uptake. They find that testosterone induces differential gene expression with a focus on metabolic enzymes. Specifically, they identify increased expression of enzymes that regulate cholesterol and fatty acid synthesis, leading to increased production of 18:1 oleic acid.
Strength:
Oleic acid is secreted by seminal vesicle epithelial cells and taken up by sperm, inducing an increase in mitochondrial respiration. The difference in sperm motility and in vivo fertilization in the presence of 18:1 oleic acid and the absence of testosterone is small but significant, suggesting that the authors have identified one of the fertilization-supporting factors in seminal plasma.
Weaknesses:
Further studies are required to investigate the effect of other seminal vesicle components on sperm capacitation to support the author's conclusions. The author's experiments focused on potential testosterone-induced changes in the rate of seminal vesicle epithelial cell glycolysis and oxphos, however, provide conflicting results and a potential correlation with seminal vesicle epithelial cell proliferation should be confirmed by additional experiments.
-
Reviewer #3 (Public Review):
Summary:
Male fertility depends on both sperm and seminal plasma, but the functional effect of seminal plasma on sperm has been relatively understudied. The authors investigate the testosterone-dependent synthesis of seminal plasma and identify oleic acid as a key factor in enhancing sperm fertility.
Strengths:
The evidence for changes in cell proliferation and metabolism of seminal vesicle epithelial cells and the identification of oleic acid as a key factor in seminal plasma is solid.
Weaknesses:
The evidence that oleic acids enhance sperm fertility in vivo needs more experimental support, as the main phenotypic effect in vitro provided by the authors remains simply as an increase in the linearity of sperm motility, which does not necessarily correlate with enhanced sperm fertility.
-
