Wound-Induced Syncytia Outpace Mononucleate Neighbors during Drosophila Wound Repair
Curation statements for this article:-
Curated by eLife

eLife Assessment
This important work addresses a very relevant biological question: what is the cellular basis of wound healing? Using the Drosophila pupal notum as a model, the paper provides an elegant, thorough, descriptive characterization of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors meticulously characterize the cell-cell fusion events during wound healing and inhibit cell fusion to show to that it is necessary to speed wound closure. In addition, the study provides convincing evidence that cell fusion allows actin resources at be partitioned to the leading edge.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
In response to injury, cells proliferate, migrate and invade to replace missing cells and close wounds. However, the role of other wound-induced cell behaviors is not understood, including the formation of syncytia (multinucleated cells). Here, we use in vivo live imaging to analyze wound-induced syncytia in mitotically competent Drosophila pupae. We find that almost half the epithelial cells near a wound fuse to form large syncytia. When the autophagy gene Atg1 is knocked down, fewer syncytia form, and wounds close more slowly. Further, a computational model of tissue fluidity indicates that cell fusion speeds wound closure time by about one third. Syncytia use several routes to speed wound repair: they outpace diploid cells at the wound margin to lead the initial resealing of the wound; they reduce the need for intercalation as the tissue reshapes during closure; and they pool resources of their component cells to concentrate them toward the wound margin. In addition to wound healing, these properties of syncytia are likely to contribute to their roles in development and pathology.
Article activity feed
-

-
-

eLife Assessment
This important work addresses a very relevant biological question: what is the cellular basis of wound healing? Using the Drosophila pupal notum as a model, the paper provides an elegant, thorough, descriptive characterization of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors meticulously characterize the cell-cell fusion events during wound healing and inhibit cell fusion to show to that it is necessary to speed wound closure. In addition, the study provides convincing evidence that cell fusion allows actin resources at be partitioned to the leading edge.
-
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental yet poorly understood biological process.
Comments on revised version.
The manuscript overall is significantly improved and authors addressed …
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental yet poorly understood biological process.
Comments on revised version.
The manuscript overall is significantly improved and authors addressed majority of my concerns. The addition of the computational vertex model (Figure 7) as well as Atg1 RNAi (Figure 4) to inhibit cell fusion provide more mechanistic insight to their study. However, the analysis of Atg1 RNAi wound assay falls short as it does directly measure changes in syncytium frequency nor size to confirm that cell fusion is reduced. The authors should quantify the number of nuclei per syncytium over the 2hr wound healing period as performed for WT in Figure 1C. It would have been ideal if they could have also performed the Act-GFP spreading assay in WT and Atg1 RNAi strains to determine if Act-GFP movement is dependent on cell fusion as purposed. At the least, further quantification of Atg1 RNAi phenotype is warranted to support their conclusions.
-
Reviewer #2 (Public Review):
Summary:
Overall, this study provides a thorough description of the formation of syncytia following wounding of the proliferation-competent diploid epithelium of the pupal notum. While this phenomenon has already been described briefly for this particular tissue by the Galko lab in Wang et al 2015, the authors provide a much more detailed description and characterisation of the process providing some novel insights (radial versus tangential border breakdown, cell shrinkage, timings, syncytia outcompeting mononucleated cells, etc.).
Strengths:
This paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors show that laser-induced wounding of this diploid, proliferation-competent epithelium …
Reviewer #2 (Public Review):
Summary:
Overall, this study provides a thorough description of the formation of syncytia following wounding of the proliferation-competent diploid epithelium of the pupal notum. While this phenomenon has already been described briefly for this particular tissue by the Galko lab in Wang et al 2015, the authors provide a much more detailed description and characterisation of the process providing some novel insights (radial versus tangential border breakdown, cell shrinkage, timings, syncytia outcompeting mononucleated cells, etc.).
Strengths:
This paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors show that laser-induced wounding of this diploid, proliferation-competent epithelium results in the formation of syncytia of various sizes in the first few cell rows around the wound edge, which progressively become bigger as healing proceeds. This results in ~50% of cells becoming part of these syncytia. The cell fusion events were convincingly demonstrated by showing the disappearance of p120ctnRFP and E-Cadherin-GFP from cell-cell borders as well as cytoplasmic GFP mixing of GFP-positive cells with a GFP-negative cell.
Apart from cell-cell fusion by border breakdown that mostly happens in the first 2h following wounding, the authors also found that at later stages of wound healing cell shrinkage following cytoplasmic mixing contributed to syncytia formation.
Next, the authors provided some convincing evidence that syncytia outcompete mononuclear cells for being positioned in the first cell row around the wound.
The authors then show that radial border breakdown occurs much less frequently than tangential border breakdown. They suggest that radial border breakdown reduces the requirement for cell-cell intercalations. They also hypothesise that tangential border breakdown might allow fused cells to share resources and provide more resources to be used near the wound edge, e.g. for actomyosin cable formation. To test this, the authors generate single-cell clones that overexpress Actin-GFP. They then show convincingly how a single Actin-GFP-positive cell in the second cell row fuses with one GFP-negative cell in the first cell row. The Actin-GFP signal then spreads in the fused cell and labels some previously unlabelled actin-rich structure near the wound edge which most likely is the actomyosin cable. This provides some evidence for resource sharing by cytoplasmic mixing following fusion.
Comments on revised version:
The authors have extended their original manuscript by adding two key parts. First, they show a role of Atg1 in mediating cell fusion (Figure 4). Second, they provide additional evidence for a contribution of radial border fusions to wound closure through its effect on tissue fluidity and through computational modelling (Figure 7).
This new version of the manuscript is greatly improved and provides significant new insights into the role of syncytia in aiding wound repair. There are just a few minor, yet important, additions needed to back up Figure 4 which should not require new experiments.
Minor but important points:
The authors show a role of Atg1 in mediating syncytia formation in Figure 4. However, since the Pnr>+ side of the wound closes slower than the non-Pnr side (control side), a few additions to this figure would be important and should not require additional experiments.
(1) The authors should show, similar to the data shown in Figure 4D of the wound radius over time for control versus Pnr>Atg1RNAi, also the same type of data for control versus Pnr>+.
(2) Since Pnr>+ also slows down wound healing, albeit to a lesser extent than Pnr>Atg1, the authors should also show an extra graph that provides evidence that Pnr>Atg1RNAi reduces syncytia formation more than Pnr>+ does. E.g. Two graphs could be added that show individual cell size at 4 or 5h post wounding for control versus Pnr>Atg1RNAi as well as for control versus Pnr>+ and also another graph with the same data but comparing cell size between Pnr>+ and Pnr>Atg1RNAi. Otherwise, if the expected minimum cell size for a syncytium is easy to estimate, a graph could be added that shows the percentage of cells that are above this threshold (e.g. above 100 square micron) for control versus Pnr>Atg1RNAi and control versus Pnr>+ and Pnr>+ versus Pnr>Atg1RNAi.
-
Reviewer #3 (Public Review):
In this revised manuscript, White et al. aimed to understand the wound-induced syncytia formation behavior in wound repair of Drosophila melanogaster pupal notum. For this purpose, the authors characterized two different types of adherens junctions' outcomes during syncytia formation around the wound region - border breakdown versus apical shrinking which appear to happen in different time points and for different time durations. The authors characterized cell-cell fusion events using cytoplasmic, junctional and nuclear markers. They determined that about half of the cells within 70 um radii from the wound undergo cell-cell fusion. They studied wound induction on the border between control epithelia and pnr domain suggesting that Atg1 is required for post-wound syncytia formation and wound closure. They …
Reviewer #3 (Public Review):
In this revised manuscript, White et al. aimed to understand the wound-induced syncytia formation behavior in wound repair of Drosophila melanogaster pupal notum. For this purpose, the authors characterized two different types of adherens junctions' outcomes during syncytia formation around the wound region - border breakdown versus apical shrinking which appear to happen in different time points and for different time durations. The authors characterized cell-cell fusion events using cytoplasmic, junctional and nuclear markers. They determined that about half of the cells within 70 um radii from the wound undergo cell-cell fusion. They studied wound induction on the border between control epithelia and pnr domain suggesting that Atg1 is required for post-wound syncytia formation and wound closure. They showed that during wound closure syncytia gradually invade the wound leading edge mostly by radial fusion events. The data suggests that intercalation of cells from the leading edge slows down the wound closure process. They propose that cell fluidity of syncytial cells plays a role in wound closure speed. Finally, the authors showed that actin is concentrated to the front edge of syncytia located in the wound leading edge. The authors described some aspects of syncytia formation during wound closure using different approaches. Some clarifications are needed as described below.
Major suggestions:
(1) Introduction, page 4. The examples of developmental syncytia formation of invertebrates and vertebrates are confusing. The authors may want to make the examples clear and add additional examples. Currently, readers may assume that C. elegans cell fusions occur only in the hypodermis - other structures can be mentioned like the vulva, pharyngeal muscles, glia, tail. In addition, the authors may want to add injury-induced fusions like the C. elegans' PLM and PVD neurons (Ghosh-Roy et al., 2010; Newman et al., 2015; Oren-Suissa et al., 2017).
(2) In cases where it is not clear whether fusion has occurred or whether mononucleated cells were ejected from the leading edge, membrane markers can be used. Page 6. Lines 96-99. The authors may want to use a membrane marker like RFP-PH driven by the epithelial cell promoter.
(3) Pages 8-10. The authors may want to clearly explain that apical junctions shrinking is a post fusion event. That the apical shrinking is caused by the expansion of fusion pores and the migration of apical junctions towards the basolateral domain. This is something that was clearly shown during physiological epidermal cell-cell fusion in C. elegans by Mohler et al., 1998 and 2002. A cartoon showing the process of cell-cell fusion, pore expansion and apical junction dynamics would make the manuscript much clearer.
(4) Page 9. Line 170. "...as these cells represent fusion initiation events (fusion pore) but were unable to productively stabilize and expand the site of fusion and so returned to the diploid state." The authors may want to make clear that this is an assumption that needs to be tested. Live imaging using a membrane marker may resolve whether a reversible fusion pore was generated.
(5) Page 11. It is not clear whether Atg1 is directly required for cell fusion, or that autophagy is required for efficient cell fusion or both Atg1 and autophagy participate in the fusion process.
(6) Page 12. Line 235. "Indeed, we observed that several hours after wounding, the entire leading edge was occupied by syncytia." This observation is based only on the adherens junction marker. Can they test basal cell membrane marker? Is it possible that the mononucleate cell in the leading edge is under the two syncytia?
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental, yet poorly understood biological process.
Weakn…
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental, yet poorly understood biological process.
Weaknesses:
A major weakness is that all the authors' conclusions are based on descriptive studies, in which the role of cell fusion is not directly tested. This is particularly important because other models of wound induced polyploidization have demonstrated that another cytoskeletal protein, myosin, was upregulated and dependent on endoreplication, and not cell fusion. Therefore it remains unclear to what extent cell fusion, endoreplication, or both are required to outcompete mononucleated cells as well as pool actin as described in this study.
We thank the reviewer for appreciating our live imaging and meticulous approach. In this revision we have identified that the gene Atg1 is required for wound-induced fusion in the pupal notum: when Atg1 is knocked down, there is a reduction in wound-induced cell fusions, both border breakdown and cell shrinking. Analysis of Atg1 knockdown shows that the wounds close more slowly. This is a direct test of the role of cell fusion in speeding wound closure, presented in new Fig. 4.
Reviewer #2 (Public Review):
Summary:
Overall, this study provides a thorough description of the formation of syncytia following wounding of the proliferation-competent diploid epithelium of the pupal notum. While this phenomenon has already been described briefly for this particular tissue by the Galko lab in Wang et al 2015, the authors provide a much more detailed description and characterisation of the process providing some novel insights (radial versus tangential border breakdown, cell shrinkage, timings, syncytia outcompeting mononucleated cells, etc.).
Strengths:
This paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors show that laserinduced wounding of this diploid, proliferation-competent epithelium results in the formation of syncytia of various sizes in the first few cell rows around the wound edge, which progressively become bigger as healing proceeds. This results in ~50% of cells becoming part of these syncytia. The cell fusion events were convincingly demonstrated by showing the disappearance of p120ctnRFP and E-Cadherin-GFP from cell-cell borders as well as cytoplasmic GFP mixing of GFPpositive cells with a GFP-negative cell.
Apart from cell-cell fusion by border breakdown that mostly happens in the first 2h following wounding, the authors also found that at later stages of wound healing cell shrinkage following cytoplasmic mixing contributed to sycytia formation.
Next, the authors provided some convincing evidence that syncytia outcompete mononuclear cells for being positioned in the first cell row around the wound.
The authors then show that radial border breakdown occurs much less frequently than tangential border breakdown. They suggest that radial border breakdown reduces the requirement for cell-cell intercalations. They also hypothesise that tangential border breakdown might allow fused cells to share resources and provide more resources to be used near the wound edge, e.g. for actomyosin cable formation. To test this, the authors generate single-cell clones that overexpress Actin-GFP. They then show convincingly how a single Actin-GFP-positive cell in the second cell row fuses with one GFP-negative cell in the first cell row. The Actin-GFP signal then spreads in the fused cell and labels some previously unlabelled actin-rich structure near the wound edge which most likely is the actomyosin cable. This provides some evidence for resource sharing by cytoplasmic mixing following fusion.
Weaknesses:
The authors provide some convincing evidence that syncytia outcompete mononuclear cells for being positioned in the first cell row around the wound. The authors suggest that the syncytial cells might be better able to close the wound. However, some genetic studies would need to be done to establish this more convincingly. E.g. Could the authors genetically block syncytia formation and then show that these wounds now heal slower?
We now present such data in new Fig. 4, which describes knocking down Atg1, previously shown by the Leptin lab to promote wound-induced fusions in larval epidermis. We quantify the resulting reduction in fusion in the pupal notum and show that the leading edge advances more slowly to heal the wound.
The authors suggest that radial border breakdown reduces the requirement for cell intercalation. While this might be true it also raises the question of how the various syncytia facing the wound border change shape to allow the shrinkage of the first cell row over time to allow wound closure. None of the four movies included in the study shows the whole wound healing process until the later stages, making it hard to assess this. It would be good to include one such movie showing the syncytia in the whole wound and comment on this point.
In response to the reviewer's request, we now extend Supplemental Video S1 out through 8 hours after wounding (same video as included previously but extended longer). In this video, as in many of the wounds, it is hard to determine the exact moment of closure because a syncytium extends across the wound whereas the nuclei do not. However, during the process of closure, one can clearly observe the large syncytia becoming more wedge-shaped – drastically reducing the section of their perimeter remaining in contact with the wound’s leading edge.
In addition, we now explore how syncytia reduce the need for intercalation in a computational model, presented in new Fig. 7 and Supplemental Videos S5 and S6. One can observe the modeled syncytia becoming similarly wedge-shaped. The modeling shows that the presence of syncytia and their ability to reshape can speed closure by about 1/3 even if the syncytia have no special properties aside from their relative size.
In both the experiments and models, some syncytia are also removed from the leading edge by intercalation, but the presence of syncytia reduces the total number of intercalations needed.
The authors hypothesise that tangential border breakdown might allow fused cells to share resources and provide more resources to be used near the wound edge, e.g. for actomyosin cable formation. They show convincingly through the fusion of a single Actin-GFP-positive cell in the second cell row with a GFP-negative cell in the first cell row that Actin-GFP spreads in the fused cell and labels the previously unlabelled actomyosin cable. While the hypothesis of resource sharing to improve healing is intriguing and makes sense, this experiment doesn't necessarily prove the benefit of resource sharing. It does show cytoplasmic mixing following fusion, now allowing the GFPlabelled actin to diffuse and be incorporated into the actomyosin cable. In a wild-type condition, fusion would not increase the total concentration of resources, although it would increase the total amount of resources within this bigger fused cell. The question is whether resource sharing without increasing the protein concentration is beneficial and increases the efficiency of certain wound healing mechanisms. There might be a benefit of cell fusion, if for example certain resources were only present in limited amounts or if protein transport could increase the concentration locally. To provide better evidence for the hypothesis that resource sharing improves wound healing, maybe the authors could look at the actomyosin cable in a wounded epithelium (such as in Figure 4E, F), in which all cells express MyoII-GFP. The authors could compare the average intensity of the actomyosin cable at the wound edge in mononucleated cells versus in syncytia. If resource sharing is indeed beneficial, it might be that the actomyosin cable is stronger/brighter in syncytia or it forms quicker.
We agree with the reviewer that we have not "proved the benefit of resource sharing". Because we cannot inhibit resource sharing while still allowing cell fusion, we can think of no rigorous way to test this hypothesis. We appreciate the reviewer's suggestion of quantifying the myosin at the leading edge cable, but we can imagine too many caveats to the interpretation to make it worthwhile. Rather, we accept the limitation that this is an untested, perhaps untestable, hypothesis -- but nevertheless intriguing.
We do want to clarify ideas about the concentration of resources after fusion. We agree that the overall concentration of a given resource (mass/volume) throughout a syncytium would be the same as the overall concentration in the unfused progenitor cells; however, a syncytium would have a larger total resource mass to direct subcellularly, allowing for local subcellular concentration to be greater in a syncytium vs. an unfused cell. We demonstrate this subcellular localization of actin in a syncytium twice, in Fig. 7C and E (previously Fig. 6C,E), which we think is evidence for increased local concentration.
The biggest limitation of this study is that the authors don't address how the formation of these syncytia is regulated. While the manuscript in its current form provides some valuable new insights into syncytial-driven wound closure, it would be much more informative if it also provided some mechanistic details. The authors could test if some of the mechanisms shown to regulate syncytial formation in other types of syncytia-driven wound healing are also involved here. E.g. Yorkie was shown to negatively regulate cell fusion in adult syncytial-driven wound closure (Losick et al 2013). The authors could test for the effect of Yorkie-RNAi in the epithelium on wound closure and syncytia formation. Expression of the dominant negative RacN17 also blocked cell fusion in adult syncytial-driven wound closure (Losick et al 2013).
Moreover, JNK activation was shown to be needed in larval syncytial-driven wound closure (Galko and Krasnow 2004). The authors could test JNK pathway reporters to assess pathway activation or test if the JNK pathway is needed for syncytial-driven wound closure by expressing a dominantnegative form of Basket JNK in the epithelium.
Or could syncytia formation be regulated by changes in Integrin-mediated adhesion as shown by the Galko lab in Wang et al 2015? They show that wounding provoked a striking relocalization of PINCH and ILK, indicating the disassembly of functional FA complexes concomitant with syncytium formation. Maybe the authors could investigate some of these.
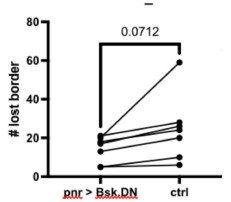
We investigated the role of JNK in fusion by expressing bskDN on one side of the wound. Comparing the numbers of border-loss fusion on each side, we did not find a significant difference in our seven-sample cohort (see Author response image 1). If we had increased the sample size, we may have found a significant difference with a small effect size, but because of the small difference in fusions on each side we did not think this was worth pursuing. Instead, we include data that the autophagy gene Atg1 is required for cell fusion in new Fig. 4, which begins to address mechanism, and relates the wound-induced fusion described here in pupae to wound-induced fusion shown in larvae. A complete mechanism for wound-induced fusion is outside the scope of this paper, as we focus on the function of syncytia in healing wounds.
Author response image 1.
Another general question that the authors raise but don't address enough is whether syncytia-driven wound closure in proliferation-competent epithelia is any different from the one in post-mitotic, polyploid epithelia. Since the mechanism regulating the former is not known, this remains unclear.
We now include a paragraph on this question in the discussion.
Finally, it is not clear, whether syncytia in these proliferation-competent epithelia get resolved after wound healing. Do they get removed and replaced by mononucleated proliferation-competent cells or do the syncytia stay in the epithelium like a scar? The authors should provide some images of wound areas a few hours after wound closure is complete and comment on this.
To answer the reviewer’s question: some but not all syncytia do get removed during wound closure by remarkable apoptotic/extrusion events. This will be the subject of a future manuscript, as it is outside the scope of this paper focusing on the function of syncytia in promoting wound healing.
Minor points:
Figure 3: It would be better to have the microcopy images alongside the quantifications.
The images in Figs. 1 and 2 show the border breakdown and shrinking cells, and we do not see benefit in adding them in Fig. 3.
Figure 4A: The syncytium at the wound edge here doesn't look straight but wavy. Does it not form an actomyosin cable that straightens the front? Or are there lamellipodia/filopodia?
We assume the reviewer is asking about the wavy edge outlined at 400 min after wounding (now Fig. 5A). As shown by Jacinto and colleagues in the first pupal wounding paper (JCB 2013), the actin cable forms quickly, within 15 minutes; much later actin protrusions extend from the leading edge to close the wound. This result is consistent with the wavy edge 400 min after wounding.
248: The authors suggest an interesting hypothesis that mitochondria or ER could be pooled in fused cells. It would be nice to see some evidence: e.g. by labeling mitochondria and assessing where they are in syncytia versus mononucleated cells and whether they are concentrated around the wound edge.
Although we don't think that exploring mitochondria or ER is central to this manuscript, we agree it would be an interesting question for the future.
141-145 (Figure 4B and C) This example is not completely convincing. First, it is hard to see where the wound edge is. Second, it would be good to include an even later time point when the cell is clearly no longer at the wound edge.
We have revised this figure, now Fig. 5B,C, to include a later image at 360 min after wounding healing, and this additional panel clarifies that the smaller cell leaves the wound edge. As noted in the text, the wound edge is indicated by the cell borders lacking p120ctn.
Reviewer #3 (Public Review):
Summary:
White et al. described laser-induced wound healing of the Drosophila pupal notum. They found that the epithelial monolayer is dynamically induced to form syncytia by cell-cell fusion as an important part of repair. They reveal two processes: cell shrinking and border breakage that occur as part of syncytia formation. Expression of GFP in the cytoplasms of some epithelial cells reveals that cytoplasmic contents mix following injury and the GFP rapidly diffuses between cells. Using live imaging they observe that syncytia expand towards the wound, maintain their positions close to the leading edge, and apparently displace smaller cells. They propose that syncytia redistribute cellular components towards the wound facilitating repair and show that labelled actin becomes concentrated at the leading edge.
Strengths:
The manuscript is interesting and on an important and emerging topic of wound healing in a genetically tractable organism. The manuscript is very well written.
Weaknesses:
There are three major issues that the authors must address: 1. Is cell-cell fusion sufficient to enhance/facilitate wound healing? 2. Characterization of "border breakdown"; Is this phenomenon disassembly of apical junctions following membrane fusion? 3. Are cells really shrinking or is it only the apical domains that "shrink" as the cells join the syncytium.
We thank the reviewer for recognizing the importance of this topic. Our responses to the specific weaknesses are below.
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
Major Components:
(1) For syncytia measurements the nuclei are labeled with histone-GFP which is expressed in all cell types. How do you know the nuclei within the cell junctions are epithelial and not another cell type, such as immune cells recruited to the injury site? It would be helpful to verify the number of nuclei per cell using an epithelial-specific nuclear marker as well. This could be via epithelial Gal4-specific expression of a UAS-nls-GFP.
This is an interesting point. In response to the reviewer's question, we investigated by doing the converse experiment, labeling immune cells with hml-Gal4, UAS-GFP, and observing what they do after wounding (analyzing six wounded pupae). They do get recruited to the wound, but they remain either in the wound center or at the basal side of the leading edge. Because they are labeled with cytoplasmic GFP, we would be able to ascertain whether they fused with epithelial cells because they would share their GFP with epithelial cells in the epithelial plane, and they did not. Thus we are confident that the many syncytial nuclei are not derived from immune cells. Our live tracking throughout the manuscript, and specifically of GFP-labeled clones, also supports our interpretation that syncytial nuclei derive from epithelial cells.
(2) The manuscript focuses on cell fusion, but other mechanisms of cell enlargement have been observed to occur during wound healing via endoreplication. To what extent do epithelial cells in pupae notum endocycle or endomitosis post injury? It is unclear if the increase in syncytia size during a 1-2hr period could also be due to endomitosis, which would also increase nuclear number.
Since the first submission of this manuscript, we published our results demonstrating limited wound-induced endoreplication after this type of explosive laser injury to the pupal notum (White et al, 2024, PMID: 38495588). We chose to publish this work separately because we could not offer the same degree of depth for endoreplication as we could for fusion: our pupal notum injury model is extremely well-suited to analyzing cell fusion and wound closure by live imaging; however, it is not particularly well-suited for analyzing endoreplication in fixed tissue. With respect to reviewer's question about endomitosis -- i.e. nuclear divisions that are not accompanied by cell divisions -- even after many years we have not observed an endomitosis event, which would be visible by live imaging, whereas we frequently and easily observe mitosis of diploid cells.
(3) One of the major conclusions of this study is that cell fusion is necessary to pool resources at the leading edge. Therefore it is critical that authors identify a mechanism to inhibit cell fusion to test this assumption.
We now include new Fig. 4, an analysis of the role of Atg1 in promoting wound-induced fusion and wound closure. These results build on the finding of the Leptin lab (Kakanj et al, 2022) that autophagy genes are required for fusion. Our results are consistent with the model that syncytia speed wound closure.
(4) There is evidence that myosin increases in endoreplicating cells during wound healing hence it is, maybe equally - if not more - probable that the increase in resources (here actin-GFP) at the leading edge is dependent on endoreplication instead of cell fusion.
Some of the new data we provide for this manuscript is a correlation between cell size and distance traveled, showing that larger cells travel more within the wound (Fig. 4F,G). Endoreplication would certainly be expected to contribute to increasing cell size, and our published 2024 data indicates that there can be one extra S-phase induced by these types of wounds. Doubling the genome is not a significant contribution to cell size compared to the 10s of nuclei we observe in syncytia from fusion. Nevertheless, we do not claim that actin is the only important resource that can be pooled subcelluarly for the benefit of the cell; we use it only as a proof-of-principle. Finally, we discuss the work on myosin in wound-induced endoreplicating cells (Losick and Duhaime, 2021).
Reviewer #3 (Recommendations For The Authors):
Major comments
(1) Can induction of epithelial fusion enhance wound healing?
Different epithelial cell-cell fusion processes have been well-characterized: i) Trophoblast fusion in the placenta mediated by Syncytins. ii) Viral induced cell-cell fusion mediated by diverse viral glycoproteins (e.g. gp41 from HIV, Hemaglutinin from Influenza, GP from Ebola, and G glycoprotein from VSV). iii) Epidermal, myoepithelial, and other epithelial cell-cell fusion in C. elegans mediated by EFF-1 and AFF-1. iv) Cell-cell fusion in the eye lens (unknown fusogens). The authors may want to compare and discuss the temporal dynamics and intermediates observed in the diverse processes of epithelial cell-cell fusion with the characterization of syncytia formation during wound healing of the Drosophila pupal notum. Since some of these characterized cell-cell fusogens can fuse heterologous cells, including Drosophila S2 cells (Shilagardi et al., 2013; https://pubmed.ncbi.nlm.nih.gov/23470732/), the authors may consider expressing these fusogens in Drosophila pupal notum before, during and after injury. This could determine whether syncytia formation is sufficient to stimulate efficient wound healing.
We thank the reviewer for the suggestion of comparing and discussing temporal dynamics and intermediates observed in the many types of epithelial fusion that are well understood. Regretfully, we do not think this article is the right venue for such a complex discussion, especially since we have little by way of comparison in our own wound-induced fusion data. As for overexpression of fusogens, it is an intriguing idea to force cell fusion with a heterologous fusogen such as EFF-1 and then investigate any resulting changes in wound healing. However, since half the cells within 70 µm of the wound already fuse even without a heterologous fusogen, it seems unlikely we could meaningfully increase the level of cell fusion unless we expressed the fusogen universally, forcing the fusion of nearly all the epithelial cells as well as other cells throughout the body that express pnr-Gal4. Because the overexpression of EFF-1 in C .elegans results in lethality (PMID: 26854231), a widespread induction of fusion would be expected to cause other types of physiological problems that would interfere with the interpretation of wound closure rates. Further, the conditional expression tools in Drosophila allow excellent spatial control, but temporal control is still somewhat low-resolution, so that we would have difficulty expressing EFF-1 before, during, and after wounding at times that would be relevant to understanding wound healing.
(2) The phenomenon of "border breakdowns" described here is not clear. The authors are probably studying the disassembly of the apical junctions following the initiation of membrane fusion and pore expansion. This should be clarified by using membrane labels to directly observe membrane fusion. Researchers have used electron microscopy and membrane fluorescent probes to follow cell-cell fusion. For example, GPI-mCherry, FM4-64, lipid-modified-GFPs (e.g. PH-domain fluorescently labeled proteins) DiO, DiI, and many others. See for example: Markosyan et al., 2016; https://pubmed.ncbi.nlm.nih.gov/26730950/; Mohler et al., 1998; https://pubmed.ncbi.nlm.nih.gov/9768364/; Meng et al., 2020; https://pubmed.ncbi.nlm.nih.gov/32668210/.
We agree completely with the reviewer, that border breakdowns represent the disassembly of apical junctions following initiation of membrane fusion and pore expansion. Direct evidence for this order of events is found in the video stills of Figure 1 panel I and video S2, which show that cytoplasmic GFP is transferred to the fusion partner 14 minutes before there is a visible decrease in the apical adherens junction marker p120ctn. The reproducibility of this order of events is documented in Fig. 3: among 107 GFP-labeled cells, 30 of them first visibly shared GFP with a fusion partner, and then 11/30 displayed border breakdown, 16/30 displayed cell shrinking, and 3/30 did not fuse. This last category is consistent with a fusion pore that closed rather than expanded productively. Although we have obtained TEM images of wound-induced fusion pores, these are included in another manuscript currently in revision and so cannot be included here, and further these EM images do not shed light on border breakdown per se, as only live imaging can establish the relationship between border breakdown and pore formation (GFP-sharing).
(3) The observation of cell shrinking may be misleading. The process the authors describe as "cell shrinking" may involve shrinking of the apical domain, maintaining the cell volume. To clarify this process, the authors may simultaneously label the apical and basolateral domains. It is possible that fusion pore formation occurs in the basolateral, apical, or both domains. The apical shrinking could reflect the migration of the apical junctions following fusion. A similar process has been described in epidermal and vulval cells of C. elegans and other nematodes (Mohler et al., 1998; https://pubmed.ncbi.nlm.nih.gov/9768364/; Sharma-Kishore et al., 1999; https://pubmed.ncbi.nlm.nih.gov/9895317/; Kolotuev and Podbilewicz 2008; https://pubmed.ncbi.nlm.nih.gov/18031720/).
We thank the reviewer for pointing out these examples of cell fusion in nematodes, and we now compare our findings to Mohler et al, 1998. In Fig. 2D, we specifically investigated what happened to the cell volume of these shrinking cells, and we hope we have now clarified both the text and the annotations on the figure to make our findings more clear. In the X-Z plane, the entire cell volume of two shrinking cells is visible from cytoplasmic GFP labeling. For both cells, the cytoplasmic volume moves laterally into the neighboring syncytia, appearing to initiate the movement from the basal-most area of the cell so that 150 minutes after wounding, both cells have a reduced apical footprint and only a whisp of apically-oriented cytoplasm, with the remainder of the cytoplasm having moved into the syncytia. These images make it clear that fusion is occuring, and that when the apical area disappears the corresponding cytoplasm has also moved into the territory of the neighboring syncytium. In response to the reviewer's suggestion, we did try labeling basolateral domains, but the fluorescent proteins we examined are not restricted to the basolateral domain and are difficult to interpret.
Minor comments
(1) Lines 40-43. Repair of injuries has also been observed in non-proliferative syncytial epidermal cells and involves cell-cell fusogens. The authors may want to include this reference: Meng et al., 2020; https://pubmed.ncbi.nlm.nih.gov/32668210/.
We thank the reviewer for the suggestion, and we have included this reference in the Discussion paragraph about fusogens.
(2) Lines 128-130. Is "Shrinking fusion" an "artefact"?
The apical junction shrinks not the cell. I suggest following basolateral membranes to see whether the cell is indeed shrinking as it fuses. The authors may want to share whether the cell volume is maintained but spills into an existing syncytium; the apical junction shrinks because it disappears/disassembles (see also Major comment 3).
As discussed in Major comment 3, we do provide evidence that the cell cytoplasm spills into an existing syncytium. Perhaps the reviewer finds the term "shrinking cell" to be misleading, as we all agree that the cell contents do not disappear. We have updated the manuscript to use the term "apical shrinking" throughout.
(3) Lines 157-159. Are these small cells or instead they are small apical junctions? The interpretation should include basolateral domains of the small cells to determine their size! It is also possible that some small cells have fused with the syncytia but on the basolateral domain without apical junction disassembly.
We appreciate the reviewer's rigor. As noted above, we were not able to analyze the basolateral domains of these cells. Because our all analyses are live-imaging videos, we are able to identify the cells are undergoing apical shrinking and clearly delineate those from stable diploid cells. We now realize that the term "small cells" is confusing and can be mixed up with apical shrinking. These cells are not "small" but normal sized, small only in comparison with the gigantic syncytia around them. We have removed the term "small" from this description.
(4) Lines 204-206. Many genes required for myoblast fusion in Drosophila have been shown to play a role in different stages of cell-cell fusion. Do they play roles in epithelia fusion during wound closure in the pupal notum?. For example, actin polymerization? Dynamin? Ig-domain and integrin cell adhesion machineries?
We now provide a new Fig. 4 that shows that the autophagy gene Atg1 reduces wound-induced cell fusion, as it does in larvae (Kakanj et al, 2022), and importantly these wounds close more slowly. We have not analyzed mutants in actin polymerization because we are confident they would interrupt many aspects of wound healing. The Galko lab has identified that integrins suppress wound-induced cell fusion in larval epidermis, but we have not tested these. We have a manuscript in revision demonstrating a requirement for Dynamin and other endocytosis genes in wound-induced fusion, and without dynamin-mediated fusion, these wounds close more slowly.
-
-
eLife assessment
This work addresses an important biological question: what is the cellular basis of wound healing? Using the Drosophila pupal notum as a model, the paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors meticulously characterize the cell-cell fusion events during wound healing, but without any mechanisms to inhibit cell fusion, it is incomplete, since it remains unclear whether cell fusion is required or not for speeding wound healing and/ or increasing the level of actin resources at the leading edge.
-
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental, yet poorly understood biological process.
Weaknesses:
A major weakness is that all the authors' conclusions are based on descriptive studies, in …
Reviewer #1 (Public Review):
Summary:
This study aims to understand how cell fusion contributes to wound healing using a laser-induced injury in the notum epithelium of a developing fruit fly. The authors meticulously characterize the epithelial fusion events using a live imaging approach and report that syncytia arise by 'border breakdown' and 'cell shrinking'. The syncytial epithelial cells also appear to outcompete mononucleated cells and preferentially dissolve their tangential borders, which correlates with the accumulation of actin at the leading edge.
Strengths:
The strength of this study is the authors' live imaging approach to capture these dynamic fusion events that are a fundamental, yet poorly understood biological process.
Weaknesses:
A major weakness is that all the authors' conclusions are based on descriptive studies, in which the role of cell fusion is not directly tested. This is particularly important because other models of wound-induced polyploidization have demonstrated that another cytoskeletal protein, myosin, was upregulated and dependent on endoreplication, and not cell fusion. Therefore it remains unclear to what extent cell fusion, endoreplication, or both are required to outcompete mononucleated cells as well as pool actin as described in this study.
-
Reviewer #2 (Public Review):
Summary:
Overall, this study provides a thorough description of the formation of syncytia following wounding of the proliferation-competent diploid epithelium of the pupal notum. While this phenomenon has already been described briefly for this particular tissue by the Galko lab in Wang et al 2015, the authors provide a much more detailed description and characterisation of the process providing some novel insights (radial versus tangential border breakdown, cell shrinkage, timings, syncytia outcompeting mononucleated cells, etc.).
Strengths:
This paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors show that laser-induced wounding of this diploid, proliferation-competent epithelium …
Reviewer #2 (Public Review):
Summary:
Overall, this study provides a thorough description of the formation of syncytia following wounding of the proliferation-competent diploid epithelium of the pupal notum. While this phenomenon has already been described briefly for this particular tissue by the Galko lab in Wang et al 2015, the authors provide a much more detailed description and characterisation of the process providing some novel insights (radial versus tangential border breakdown, cell shrinkage, timings, syncytia outcompeting mononucleated cells, etc.).
Strengths:
This paper provides an elegant, thorough, descriptive characterisation of syncytia-driven wound closure using state-of-the-art confocal live imaging of the pupal notum. The authors show that laser-induced wounding of this diploid, proliferation-competent epithelium results in the formation of syncytia of various sizes in the first few cell rows around the wound edge, which progressively become bigger as healing proceeds. This results in ~50% of cells becoming part of these syncytia. The cell fusion events were convincingly demonstrated by showing the disappearance of p120ctnRFP and E-Cadherin-GFP from cell-cell borders as well as cytoplasmic GFP mixing of GFP-positive cells with a GFP-negative cell.
Apart from cell-cell fusion by border breakdown that mostly happens in the first 2h following wounding, the authors also found that at later stages of wound healing cell shrinkage following cytoplasmic mixing contributed to sycytia formation.
Next, the authors provided some convincing evidence that syncytia outcompete mononuclear cells for being positioned in the first cell row around the wound.
The authors then show that radial border breakdown occurs much less frequently than tangential border breakdown. They suggest that radial border breakdown reduces the requirement for cell-cell intercalations. They also hypothesise that tangential border breakdown might allow fused cells to share resources and provide more resources to be used near the wound edge, e.g. for actomyosin cable formation. To test this, the authors generate single-cell clones that overexpress Actin-GFP. They then show convincingly how a single Actin-GFP-positive cell in the second cell row fuses with one GFP-negative cell in the first cell row. The Actin-GFP signal then spreads in the fused cell and labels some previously unlabelled actin-rich structure near the wound edge which most likely is the actomyosin cable. This provides some evidence for resource sharing by cytoplasmic mixing following fusion.
Weaknesses:
The authors provide some convincing evidence that syncytia outcompete mononuclear cells for being positioned in the first cell row around the wound. The authors suggest that the syncytial cells might be better able to close the wound. However, some genetic studies would need to be done to establish this more convincingly. E.g. Could the authors genetically block syncytia formation and then show that these wounds now heal slower?
The authors suggest that radial border breakdown reduces the requirement for cell intercalation. While this might be true it also raises the question of how the various syncytia facing the wound border change shape to allow the shrinkage of the first cell row over time to allow wound closure. None of the four movies included in the study shows the whole wound healing process until the later stages, making it hard to assess this. It would be good to include one such movie showing the syncytia in the whole wound and comment on this point.
The authors hypothesise that tangential border breakdown might allow fused cells to share resources and provide more resources to be used near the wound edge, e.g. for actomyosin cable formation. They show convincingly through the fusion of a single Actin-GFP-positive cell in the second cell row with a GFP-negative cell in the first cell row that Actin-GFP spreads in the fused cell and labels the previously unlabelled actomyosin cable. While the hypothesis of resource sharing to improve healing is intriguing and makes sense, this experiment doesn't necessarily prove the benefit of resource sharing. It does show cytoplasmic mixing following fusion, now allowing the GFP-labelled actin to diffuse and be incorporated into the actomyosin cable. In a wild-type condition, fusion would not increase the total concentration of resources, although it would increase the total amount of resources within this bigger fused cell. The question is whether resource sharing without increasing the protein concentration is beneficial and increases the efficiency of certain wound healing mechanisms. There might be a benefit of cell fusion, if for example certain resources were only present in limited amounts or if protein transport could increase the concentration locally. To provide better evidence for the hypothesis that resource sharing improves wound healing, maybe the authors could look at the actomyosin cable in a wounded epithelium (such as in Figure 4E, F), in which all cells express MyoII-GFP. The authors could compare the average intensity of the actomyosin cable at the wound edge in mononucleated cells versus in syncytia. If resource sharing is indeed beneficial, it might be that the actomyosin cable is stronger/brighter in syncytia or it forms quicker.
The biggest limitation of this study is that the authors don't address how the formation of these syncytia is regulated. While the manuscript in its current form provides some valuable new insights into syncytial-driven wound closure, it would be much more informative if it also provided some mechanistic details. The authors could test if some of the mechanisms shown to regulate syncytial formation in other types of syncytia-driven wound healing are also involved here. E.g. Yorkie was shown to negatively regulate cell fusion in adult syncytial-driven wound closure (Losick et al 2013). The authors could test for the effect of Yorkie-RNAi in the epithelium on wound closure and syncytia formation. Expression of the dominant negative RacN17 also blocked cell fusion in adult syncytial-driven wound closure (Losick et al 2013).
Moreover, JNK activation was shown to be needed in larval syncytial-driven wound closure (Galko and Krasnow 2004). The authors could test JNK pathway reporters to assess pathway activation or test if the JNK pathway is needed for syncytial-driven wound closure by expressing a dominant-negative form of Basket JNK in the epithelium.
Or could syncytia formation be regulated by changes in Integrin-mediated adhesion as shown by the Galko lab in Wang et al 2015? They show that wounding provoked a striking relocalization of PINCH and ILK, indicating the disassembly of functional FA complexes concomitant with syncytium formation. Maybe the authors could investigate some of these.
Another general question that the authors raise but don't address enough is whether syncytia-driven wound closure in proliferation-competent epithelia is any different from the one in post-mitotic, polyploid epithelia. Since the mechanism regulating the former is not known, this remains unclear.
Finally, it is not clear, whether syncytia in these proliferation-competent epithelia get resolved after wound healing. Do they get removed and replaced by mononucleated proliferation-competent cells or do the syncytia stay in the epithelium like a scar? The authors should provide some images of wound areas a few hours after wound closure is complete and comment on this.
Minor points:
Figure 3: It would be better to have the microcopy images alongside the quantifications.
Figure 4A: The syncytium at the wound edge here doesn't look straight but wavy. Does it not form an actomyosin cable that straightens the front? Or are there lamellipodia/filopodia?
248: The authors suggest an interesting hypothesis that mitochondria or ER could be pooled in fused cells. It would be nice to see some evidence: e.g. by labeling mitochondria and assessing where they are in syncytia versus mononucleated cells and whether they are concentrated around the wound edge.
141-145 (Figure 4B and C) This example is not completely convincing. First, it is hard to see where the wound edge is. Second, it would be good to include an even later time point when the cell is clearly no longer at the wound edge.
-
Reviewer #3 (Public Review):
Summary:
White et al. described laser-induced wound healing of the Drosophila pupal notum. They found that the epithelial monolayer is dynamically induced to form syncytia by cell-cell fusion as an important part of repair. They reveal two processes: cell shrinking and border breakage that occur as part of syncytia formation. Expression of GFP in the cytoplasms of some epithelial cells reveals that cytoplasmic contents mix following injury and the GFP rapidly diffuses between cells. Using live imaging they observe that syncytia expand towards the wound, maintain their positions close to the leading edge, and apparently displace smaller cells. They propose that syncytia redistribute cellular components towards the wound facilitating repair and show that labelled actin becomes concentrated at the leading edge.
Reviewer #3 (Public Review):
Summary:
White et al. described laser-induced wound healing of the Drosophila pupal notum. They found that the epithelial monolayer is dynamically induced to form syncytia by cell-cell fusion as an important part of repair. They reveal two processes: cell shrinking and border breakage that occur as part of syncytia formation. Expression of GFP in the cytoplasms of some epithelial cells reveals that cytoplasmic contents mix following injury and the GFP rapidly diffuses between cells. Using live imaging they observe that syncytia expand towards the wound, maintain their positions close to the leading edge, and apparently displace smaller cells. They propose that syncytia redistribute cellular components towards the wound facilitating repair and show that labelled actin becomes concentrated at the leading edge.
Strengths:
The manuscript is interesting and on an important and emerging topic of wound healing in a genetically tractable organism. The manuscript is very well written.
Weaknesses:
There are three major issues that the authors must address:
(1) Is cell-cell fusion sufficient to enhance/facilitate wound healing?
(2) Characterization of "border breakdown"; Is this phenomenon disassembly of apical junctions following membrane fusion?
(3) Are cells really shrinking or is it only the apical domains that "shrink" as the cells join the syncytium?
-
-
-
-
-
