The role of GABA in semantic memory and its neuroplasticity
Curation statements for this article:-
Curated by eLife

eLife Assessment
Jung et al. present valuable work on the relationship between gamma-aminobutyric acid (GABA) levels within the anterior temporal lobes (ATL) to semantic memory while accounting for inter-individual differences. They provide solid evidence suggesting that inhibitory continuous theta burst transcranial magnetic stimulation (cTBS TMS) increased GABA concentration and decreased the blood-oxygen dependent signal (BOLD) during a semantic task. The results will be of interest to researchers studying the neurobiology of semantic cognition.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
A fundamental challenge in neuroscience is understanding neural functioning and plasticity of the brain. The anterior temporal lobe (ATL) is a hub for semantic memory, which generates coherent conceptual representations. GABAergic inhibition plays a crucial role in shaping human cognition and plasticity, but it is unclear how this inhibition relates to human semantic memory and its plasticity. Here, we employed a combination of continuous theta burst stimulation (cTBS), MR spectroscopy and fMRI to investigate the role of GABA in semantic memory and its neuroplasticity. We found that inhibitory cTBS increased GABA concentrations in the ATL and reduced blood-oxygen level-dependent (BOLD) activation during semantic tasks. Crucially, changes in GABA were tightly linked to changes in regional activity, suggesting that GABA mediates cTBS-induced plasticity. Individuals with better semantic performance exhibited selective activity in the ATL, attributable to higher GABA levels, which can sharpen distributed semantic representations. Our results revealed a non-linear, inverted-U-shape relationship between GABA levels in the ATL and semantic performance, thus offering an explanation for the individual differences in semantic memory function and neuromodulation outcomes. These findings offer a neurochemical explanation for individual variability in neuromodulation and provide insights for developing targeted interventions for semantic impairments.
Article activity feed
-

-
-
-

eLife Assessment
Jung et al. present valuable work on the relationship between gamma-aminobutyric acid (GABA) levels within the anterior temporal lobes (ATL) to semantic memory while accounting for inter-individual differences. They provide solid evidence suggesting that inhibitory continuous theta burst transcranial magnetic stimulation (cTBS TMS) increased GABA concentration and decreased the blood-oxygen dependent signal (BOLD) during a semantic task. The results will be of interest to researchers studying the neurobiology of semantic cognition.
-
Reviewer #1 (Public review):
This study presents valuable findings on the GABA and BOLD changes induced by continuous theta burst stimulation (cTBS) and on the relationships between ATL GABA level and performance in a semantic task. However, I'm afraid that the current results are incomplete to support some primary claims of the paper, for example, the purported inverted-U-shaped relationship between GABA levels in the ATL and semantic task performance. The influence of practice effects also complicates the interpretation of the results. Additional concerns include potential double dipping in the analysis depicted in Figure 3A and the use of inconsistent behavioral measures (IE and accuracy) across various analyses.
The authors have made two beneficial revisions in this round: (1) acknowledging the insufficient data points supporting …
Reviewer #1 (Public review):
This study presents valuable findings on the GABA and BOLD changes induced by continuous theta burst stimulation (cTBS) and on the relationships between ATL GABA level and performance in a semantic task. However, I'm afraid that the current results are incomplete to support some primary claims of the paper, for example, the purported inverted-U-shaped relationship between GABA levels in the ATL and semantic task performance. The influence of practice effects also complicates the interpretation of the results. Additional concerns include potential double dipping in the analysis depicted in Figure 3A and the use of inconsistent behavioral measures (IE and accuracy) across various analyses.
The authors have made two beneficial revisions in this round: (1) acknowledging the insufficient data points supporting the inverted U-shaped curve; (2) attempting to control for practice effects. However, I believe unresolved issues remain:
(1) The authors have not addressed my specific concern about Figure 4D - the analysis attempts to fit an inverted U-shaped curve to the data without distinguishing between data points influenced by practice effects and those unaffected, rendering its reliability questionable.
(2) The authors appear to have misunderstood my question regarding Figure 3A. This issue is unrelated to practice effects. My point was that even if we randomly generated pre- and post-test data points and grouped/analyzed them according to the authors' methodology, we would still likely reproduce the pattern in Figure 3A due to the double dipping problem. Thus, this statistical analysis and its conclusions currently lack methodological validity.
(3) Regarding the inconsistency in behavioral measures, the authors' explanation fails to remove my concerns. If the authors argue that accuracy is the most appropriate behavioral dependent variable for this study, why did they employ inverse efficiency in some of their analyses? My understanding is that a study should either consistently use the single most suitable measure or report multiple measures while providing adequate discussion of inconsistent results.
-
Reviewer #3 (Public review):
As a result of a number of rounds of reviews and consultations between reviewers, Jung et al. present important work on the relationship between gamma-aminobutyric acid (GABA) levels within the anterior temporal lobes (ATL) to semantic memory while accounting for inter-individual differences. They provide solid evidence suggesting that inhibitory continuous theta burst transcranial magnetic stimulation (cTBS TMS) increased GABA concentration and decreased the blood-oxygen dependent signal (BOLD) during a semantic task.
The authors fully addressed my comments from the first and second rounds of reviews, and I do not have additional concerns. I have, however, scaled down my short assessment, given the concerns of reviewers 1 and 2.
-
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
This study examined the changes in ATL GABA levels induced by cTBS and its relationship with BOLD signal changes and performance in a semantic task. The findings suggest that the increase in ATL GABA levels induced by cTBS is associated with a decrease in BOLD signal. The relationship between ATL GABA levels and semantic task performance is nonlinear, and more specifically, the authors propose that the relationship is an inverted U-shaped relationship.
Strengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.
Weak…
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
This study examined the changes in ATL GABA levels induced by cTBS and its relationship with BOLD signal changes and performance in a semantic task. The findings suggest that the increase in ATL GABA levels induced by cTBS is associated with a decrease in BOLD signal. The relationship between ATL GABA levels and semantic task performance is nonlinear, and more specifically, the authors propose that the relationship is an inverted U-shaped relationship.
Strengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.
Weakness:
I am pleased to see the authors' feedback on my previous questions and suggestions, and I believe the additional data analysis they have added is helpful. Here are my reserved concerns and newly discovered issues.
(1) Regarding the Inverted U-Shaped Curve In the revised manuscript, the authors have accepted some of my suggestions and conducted further analysis, which is now presented in Figure 3B. These results provide partial support for the authors' hypothesis. However, I still believe that the data from this study hardly convincingly support an inverted U-shaped distribution relationship.
The authors stated in their response, "it is challenging to determine the optimal level of ATL GABA," but I think this is achievable. From Figures 4C and 4D, the ATL GABA levels corresponding to the peak of the inverted U-shaped curve fall between 85 and 90. In my understanding, this can be considered as the optimal level of ATL GABA estimated based on the existing data and the inverted U-shaped curve relationship. However, in the latter half of the inverted U-shaped curve, there are quite few data points, and such a small number of data points hardly provides reliable support for the quantitative relationship in the latter half of the curve. I suggest that the authors should at least explicitly acknowledge this and be cautious in drawing conclusions. I also suggest that the authors consider fitting the data with more types of non-linear relationships, such as a ceiling effect (a combination of a slope and a horizontal line), or a logarithmic curve.
We appreciate R1’s comments. Inverted U-shaped relationships are well-established in neuroscience, particularly in the context of neurotransmitter concentrations (e.g., dopamine, acetylcholine, noradrenaline) and their influence on cognitive functions such as working memory and cognitive control (Aston-Jones & Cohen., 2005; Cools & D'Esposito., 2011; Vijayraghavan et al., 2007; He & Zempel., 2013). Recently, Ferri et al. (2017) demonstrated an inverted U-shaped relationship between excitation-inhibition balance (EIB: the ratio of Glx and GABA) and multisensory integration, showing that both excessive and insufficient inhibition negatively impact functionality. Given that GABA is the brain’s primary inhibitory neurotransmitter, our findings suggest that ATL GABA may play a similar regulatory role in semantic memory function.
While our statistical modelling approach demonstrated that the inverted U-shaped function was the best-fitting model for our current data in explaining the relationship between ATL GABA and semantic memory, we acknowledge the limitation of having fewer data points in the latter half (right side) of the curve, where excessive ATL GABA levels are associated with poorer semantic performance. Following R1’s suggestion, we have explicitly acknowledged this limitation in the revised manuscript and exercised caution in our discussion.
Discussion, p.17, line 408
"However, our findings should be interpreted with caution due to the limitation of having fewer data points in the latter half (right side) of the inverted U-shaped curve. Future studies incorporating GABA agonists could help further validate and refine these findings."
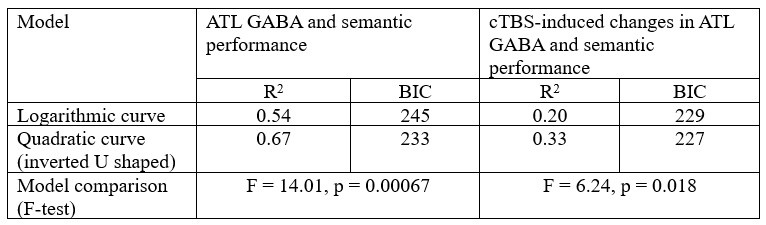
Following R1’s latter suggestion, we tested a logarithmic curve model. The results showed significant relationships between ATL GABA and semantic performance (R2 = 0.544, p < 0.001) and between cTBS-induced changes in ATL GABA and semantic performance (R2 = 0.202, p < 0.001). However, the quadratic (inverted U-shaped) model explained more variance than the logarithmic model, as indicated by a higher R2 and lower BIC. Model comparisons further confirmed that the inverted U-shaped model provided the best fit for both ATL GABA in relation to semantic performance (Fig. 4C) and cTBS-induced ATL GABA changes in relation to semantic function (Fig. 4D).
Author response table 1.
(2) In Figure 2F, the authors demonstrated a strong practice effect in this study, which to some extent offsets the decrease in behavioral performance caused by cTBS. Therefore, I recommend that the authors give sufficient consideration to the practice effect in the data analysis.
One issue is the impact of the practice effect on the classification of responders and non-responders. Currently, most participants are classified as non-responders, suggesting that the majority of the population may not respond to the cTBS used in this study. This greatly challenges the generalizability of the experimental conclusions. However, the emergence of so many non-responders is likely due to the prominent practice effect, which offsets part of the experimental effect. If the practice effect is excluded, the number of responders may increase. The authors might estimate the practice effect based on the vertex simulation condition and reclassify participants after excluding the influence of the practice effect.
Another issue is that considering the significant practice effect, the analysis in Figure 4D, which mixes pre- and post-test data, may not be reliable.
We appreciate Reviewer 1’s thoughtful comments regarding the practice effect and its potential impact on our findings. Our previous analysis revealed a strong practice effect on reaction time (RT), with participants performing tasks faster in the POST session, regardless of task condition (Fig. S3). Given our hypothesis that inhibitory ATL cTBS would disrupt semantic task performance, we accounted for this by using inverse efficiency (IE), which combines accuracy and RT. This analysis demonstrated that ATL cTBS disrupted semantic task performance compared to both control stimulation (vertex) and control tasks, despite the practice effect (i.e., faster RT in the POST session), thereby supporting our hypothesis. These findings may suggest that the effects of ATL cTBS were more subtly reflected in semantic task accuracy rather than RT.
Regarding inter-individual variability in response to rTMS/TBS, prior studies have shown that 50–70% of participants are non-responders, either do not respond or respond in an unexpected manner (Goldsworthy et al., 2014; Hamada et al., 2013; Hinder et al., 2014; Lopez-Alonso et al., 2014; Maeda et al., 2000a; Müller-Dahlhaus et al., 2008). Our previous study (Jung et al., 2022) using the same semantic task and cTBS protocol was the first to explore TBS-responsiveness variability in semantic memory, where 12 out of 20 participants (60%) were classified as responders. The proportion of responders and non-responders in the current study aligns with previous findings, suggesting that this variability is expected in TBS research.
However, we acknowledge R1’s concern that the strong practice effect may have influenced responder classification. To address this, we estimated the practice effect using the vertex stimulation condition and reclassified participants accordingly by adjusting ATL stimulation performance (IE) relative to vertex stimulation performance (IE). This reclassification identified nine responders (an increase of two), aligning with the typical responder proportion (52%) reported in the TBS literature. Overall, we replicated the previous findings with improved statistical robustness.
A 2×2×2 ANOVA was conducted with task (semantic vs. control) and session (PRE vs. POST) as within-subject factors, and group (responders vs. non-responders) as a between-subject factor. The analysis revealed a significant interaction between the session and group (F1, 15 = 10.367, p = 0.006), a marginally significant interaction between the session and task (F1, 15 = 4.370, p = 0.054), and a significant 3-way interaction between the session, task, and group (F1, 15 = 7.580, p = 0.015). Post hoc t-tests showed a significant group difference in semantic task performance following ATL stimulation (t = 2.349, p = 0.033). Post hoc paired t-test demonstrated that responders exhibited poorer semantic task performance following the ATL cTBS (t = -5.281, p < 0.001), whereas non-responders showed a significant improvement (t = 3.206, p = 0.007) (see Figure. 3A).
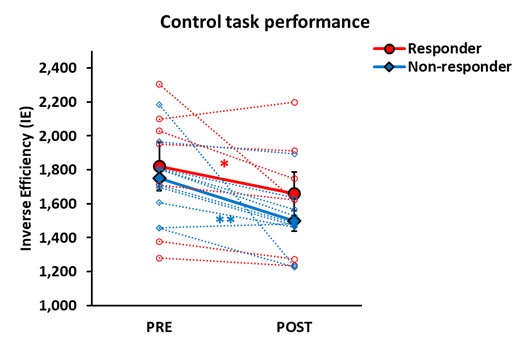
Notably, no differences were observed between responders and non-responders in the control task performance across pre- and post-stimulation sessions, confirming that the practice effect was successfully controlled (Figure. 3B).
We performed a 2 x 2 ANOVA with session (pre vs. post) as a within subject factor and with group (responders vs. non-responders) as a between subject factor to examine the effects of group in ATL GABA levels. The results revealed a significant main effect of session (F1, 14 = 39.906, p < 0.001) and group (F1, 14 = 9.677, p = 0.008). Post hoc paired t-tests on ATL GABA levels showed a significant increase in regional ATL GABA levels following ATL stimulation for both responders (t = -3.885, p = 0.002) and non-responders (t = -4.831, p = 0.001). Furthermore, we replicated our previous finding that baseline GABA levels were significantly higher in responders compared to non-responders (t = 2.816, p = 0.007) (Figure. 3C). This pattern persisted in the post-stimulation session (t = 2.555, p = 0.011) (Figure. 3C).
Accordingly, we have revised the Methods and Materials (p 26, line 619), Results (p11, line 233-261), and Figure 3.
(3) The analysis in Figure 3A has a double dipping issue. Suppose we generate 100 pairs of random numbers as pre- and post-test scores, and then group the data based on whether the scores decrease or increase; the pre-test scores of the group with decreased scores will have a very high probability of being higher than those of the group with increased scores. Therefore, the findings in Figure 3A seem to be meaningless.
Yes, we agreed with R1’s comments. However, Figure 3A illustrates interindividual responsiveness patterns, while Figure 3B demonstrates that these results account for practice effects, incorporating new analyses.
(4) The authors use IE as a behavioral measure in some analyses and use accuracy in others. I recommend that the authors adopt a consistent behavioral measure.
We appreciate Reviewer 1’s suggestion. In examining the relationship between ATL GABA and semantic task performance, we have found that only semantic accuracy—not reaction time (RT) or inverse efficiency (IE)—shows a significant positive correlation and regression with ATL GABA levels and semantic task-induced ATL activation, both in our previous study (Jung et al., 2017) and in the current study. ATL GABA levels were not correlated with semantic RT (Jung et al., 2017: r = 0.34, p = 0.14, current study: r = 0.26, p = 0.14). It should be noted that there were no significant correlations between ATL GABA levels and semantic inverse efficiency (IE) in both studies (Jung et al., 2017: r = 0.13, p = 0.62, current study: r = 0.22, p = 0.44). As a result, we found no significant linear and non-linear relationship between ATL GABA levels and RT (linear function R2 = 0.21, p =0.45, quadratic function: R2 = 0.17, p = 0.21) and between ATL GABA levels and IE (linear function R2 = 0.24, p =0.07, quadratic function: R2 = 2.24, p = 0.12).
The absence of a meaningful relationship between ATL GABA and semantic RT or IE may be due to the following reasons: 1) RT is primarily associated with premotor and motor activation during semantic processing rather than ATL activation; 2) ATL GABA is likely to play a key role in refining distributed semantic representations through lateral inhibition, which sharpens the activated representation (Jung et al., 2017; Liu et al. 2011; Isaacson & Scanziani., 2011). This sharpening process may contribute to more accurate semantic performance (Jung et al., 2017). In our semantic task, for example, when encountering a camel (Fig. 1B), multiple semantic features (e.g., animal, brown, desert, sand, etc.) are activated. To correctly identify the most relevant concept (cactus), irrelevant associations (tree) must be suppressed—a process that likely relies on inhibitory mechanisms. Given this theoretical framework, we have used accuracy as the primary measure of semantic performance to elucidate the ATL GABA function.
Reviewer #2 (Public review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data with their previous MRS study suggest an inverted u-shape between GABA concentration and behavioral performance. These results help to better understand the neuromodulatory effects of non-invasive brain stimulation on task performance.
Strengths:
Multimodal assessment of neurostimulation effects on the behavioral, neurochemical, and neural levels. In particular, the link between GABA modulation and behavior is timely and potentially interesting.
Weaknesses:
The analyses are not sound. Some of the effects are very weak and not all conclusions are supported by the data since some of the comparisons are not justified. There is some redundancy with a previous paper by the same authors, so the novelty and contribution to the field are overall limited. A network approach might help here.
Reviewer #3 (Public review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on the semantic task. This is in line with their second prediction. Finally, the authors conducted additional analyses to explore the mechanistic link between ATL inhibitory GABAergic action and semantic task performance. They show that this link is best captured by an inverted U-shaped function as a result of a quadratic linear regression model. Fitting this model to their data indicates that increasing GABA levels led to better task performance as long as they were not excessively low or excessively high. This was first tested as a relationship between GABA levels in the ATL and semantic task performance; then the same analyses were performed on the pre and post-cTBS TMS stimulation data, showing the same pattern. These results are in line with the conclusions of the authors.
Comments on revisions:
The authors have comprehensively addressed my comments from the first round of review, and I consider most of their answers and the steps they have taken satisfactorily. Their insights prompted me to reflect further on my own knowledge and thinking regarding the ATL function.
I do, however, have an additional and hopefully constructive comment regarding the point made about the study focusing on the left instead of bilateral ATL. I appreciate the methodological complexities and the pragmatic reasons underlying this decision. Nevertheless, briefly incorporating the justification for this decision into the manuscript would have been beneficial for clarity and completeness. The presented argument follows an interesting logic; however, despite strong previous evidence supporting it, the approach remains based on an assumption. Given that the authors now provide the group-level fMRI results captured more comprehensively in Supplementary Figure 2, where the bilateral pattern of fMRI activation can be observed in the current data, the authors could have strengthened their argument by asserting that the activation related to the given semantic association task in this data was bilateral. This would imply that the TMS effects and associated changes in GABA should be similar for both sites. Furthermore, it is worth noting the approach taken by Pobric et al. (2007, PNAS), who stimulated a site located 10 mm posterior to the tip of the left temporal pole along the middle temporal gyrus (MTG) and not the bilateral ATL.
We appreciate the reviewer’s constructive comment regarding the focus on the left ATL rather than bilateral ATL in our study. Accordingly, we have added the following paragraph in the Supplementary Information.
“Justification of target site selection and cTBS effects
Evidence suggests that bilateral ATL systems contribute to semantic representation (for a review, see Lambon Ralph., 2017). Consistent with this, our semantic task induced bilateral ATL activation (Fig. S2). Thus, stimulating both left and right ATL could provide a more comprehensive understanding of cTBS effects and its GABAergic function.
Previous rTMS studies have applied inhibitory stimulation to the left vs. right ATL, demonstrating that stimulation at either site significantly disrupted semantic task performance (Pobric et al., 2007, PNAS; Pobric et al., 2010, Neuropsychologia; Lambon Ralph et al., 2009, Cerebral Cortex). Importantly, these studies reported no significant difference in rTMS effects between left and right ATL stimulation, suggesting that stimulating either hemisphere produces comparable effects on semantic processing. In the current study, we combined cTBS with multimodal imaging to investigate its effects on the ATL. Given our study design constraints (including the need for a control site, control task, and control stimulation) and limitations in scanning time, we selected the left ATL as the target region. This choice also aligned with the MRS voxel placement used in our previous study (Jung et al., 2017), allowing us to combine datasets and further investigate GABAergic function in the ATL. Accordingly, cTBS was applied to the peak coordinate of the left ventromedial ATL (MNI -36, -15, -30) as identified by previous fMRI studies (Binney et al., 2010; Visser et al., 2012).
Given that TMS pulses typically penetrate 2–4 cm, we acknowledge the challenge of reaching deeper ventromedial ATL regions. However, our findings indicate that cTBS effectively modulated ATL function, as evidenced by reduced task-induced regional activity, increased ATL GABA concentrations, and poorer semantic performance, confirming that TMS pulses successfully influenced the target region. To further validate these effects, we conducted an ROI analysis centred on the ventromedial ATL (MNI -36, -15, -30), which revealed a significant reduction in ATL activity during semantic processing following ATL stimulation (t = -2.43, p = 0.014) (Fig. S7). This confirms that cTBS successfully modulated ATL activity at the intended target coordinate.”
We appreciate R3's comment regarding the approach taken by Pobric et al. (2007, PNAS), who stimulated a site 10 mm posterior to the tip of the left temporal pole along the middle temporal gyrus (MTG). This approach has been explicitly discussed in our previous papers and reviews (e.g., Lambon Ralph, 2014, Proc. Royal Society B). Our earlier use of lateral ATL stimulation at this location (Pobric et al. 2007; Lambon Ralph et al. 2009; Pobric et al. 2010) was based on its alignment with the broader ATL region commonly atrophied in semantic dementia (cf. Binney et al., 2010 for a direct comparison of SD atrophy, fMRI data and the TMS region). Since these original ATL TMS investigations, a series of distortion-corrected or distortion-avoiding fMRI studies (e.g., Binney et al 2010; Visser et al, various, Hoffman et al., various; Jackson et al., 2015) have demonstrated graded activation differences across the ATL. While weaker activation is present at the original lateral ATL (MTG) stimulation site, the peak activation is maximal in the ventromedial ATL—a finding that was also observed in the current study. Accordingly, we selected the ventromedial ATL as our target site for stimulation.
Following these points, we have revised the manuscript in the Methods and Materials.
Transcranial magnetic stimulation p23, line 525-532,
“Previous rTMS studies targeted a lateral ATL site 10 mm posterior to the temporal pole on the middle temporal gyrus (MTG) (Pobric et al. 2007; Lambon Ralph et al. 2009; Pobric et al. 2010), aligning with the broader ATL region typically atrophied in semantic dementia (Binney et al. 2010). However, distortion-corrected fMRI studies (Binney et al. 2010; Visser et al. 2012) have revealed graded activation differences across the ATL, with peak activation in the ventromedial ATL. Based on these findings, we selected the target site in the left ATL (MNI -36, -15, -30) from a prior distortion-corrected fMRI study (Binney et al. 2010; Visser et al. 2012 that employed the same tasks as our study (for further details, see the Supplementary Information).”
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
The authors have responded to all my comments and I found most of the responses reasonable and sufficient. However, I have one remaining point: I pointed out before that the scope of this paper is somehow narrow and asked for a network analysis. I found the response to my question somehow puzzling since the authors write:
"However, it is important to note that we did not find any significant correlations between ATL GABA changes and cTBS-induced changes in the functional connectivity. Consequently, we are currently preparing another paper that specifically addresses the network-level changes induced by ATL cTBS."
I don't understand the logic here. Even in the absence of significant correlations between ATL GABA changes and cTBS-induced changes in connectivity, it would be interesting to know how baseline connectivity is correlated with the induced changes. I am not sure if it is adequate to squeeze another paper out of the dataset instead of reporting it here as suggested.
We apologise that our previous response was not clear. To examine cTBS-induced network-level changes, we conducted ROI analyses targeting key semantic regions, including the bilateral ATL, inferior frontal gyrus (IFG), and posterior middle temporal gyrus (pMTG), as well as Psychophysiological Interactions (PPI) using the left ATL as a seed region. The ROI analysis revealed that ATL stimulation significantly decreased task-induced activity in the left ATL (target region) while increasing activity in the right ATL and left IFG. PPI analyses showed that ATL stimulation enhanced connectivity between the left ATL and the right ATL (both ventromedial and lateral ATL), bilateral IFG, and bilateral pMTG, suggesting that ATL stimulation modulates a bilateral semantic network.
Building on these findings, we conducted Dynamic Causal Modeling (DCM) to estimate and infer interactions among predefined brain regions across different experimental conditions (Friston et al., 2003). The bilateral ventromedial ATL, lateral ATL, IFG, and pMTG were defined as network nodes with mutual connections. Our model examined cTBS effects at the left ATL under both baseline (intrinsic) and semantic task (modulatory) conditions, estimating 56 intrinsic parameters for baseline connectivity and testing 16 different modulatory models to assess cTBS-induced connectivity changes during semantic processing. Here, we briefly summarize the key DCM analysis results: 1) ATL cTBS significantly altered effective connectivity between the left and right lateral and ventromedial ATL in both intrinsic and modulatory conditions; 2) cTBS increased modulatory connectivity from the right to the left ATL compared to vertex stimulation.
Given the complexity and depth of these findings, we believe that a dedicated paper focusing on the network-level effects of ATL cTBS is necessary to provide a more comprehensive and detailed analysis, which extends beyond the scope of the current study. It should be noted that no significant relationship was found between ATL GABA levels and ATL connectivity in both PPI and DCM analyses.
Reviewer #3 (Recommendations for the authors):
In response to my comment about the ATL activation being rather medial in the fMRI data and my concern about the TMS pulse perhaps not reaching this site, the authors offer an excellent solution to demonstrate TMS effects to such a medial ATL coordinate. I think that the analyses and figures they provide as a response to this comment and a brief explanation of this result should be incorporated into supplementary materials for methodologically oriented readers. Also, perhaps it would be beneficial to discuss that the effect of TMS on vATL remains a matter of further research to see not just if but also how TMS pulse reaches target coordinates, given the problematic anatomical location of the region.
We appreciate R3’s suggestion. Please, see our reply above.
-

-
-
eLife Assessment
This is a valuable paper that might contribute new insight into the role of GABA in semantic memory, which is a significant question in higher cognition. However, the empirical support for the main claims is incomplete. These results, once further strengthened and more appropriately discussed, will be of interest to broad readers of the neuroscience and cognitive neuroscience community.
-
Reviewer #1 (Public review):
Summary:
This study examined the changes in ATL GABA levels induced by cTBS and its relationship with BOLD signal changes and performance in a semantic task. The findings suggest that the increase in ATL GABA levels induced by cTBS is associated with a decrease in BOLD signal. The relationship between ATL GABA levels and semantic task performance is nonlinear, and more specifically, the authors propose that the relationship is an inverted U-shaped relationship.
Strengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.
Weakness:
I am pleased to see the authors' feedback on my previous questions and suggestions, and I …
Reviewer #1 (Public review):
Summary:
This study examined the changes in ATL GABA levels induced by cTBS and its relationship with BOLD signal changes and performance in a semantic task. The findings suggest that the increase in ATL GABA levels induced by cTBS is associated with a decrease in BOLD signal. The relationship between ATL GABA levels and semantic task performance is nonlinear, and more specifically, the authors propose that the relationship is an inverted U-shaped relationship.
Strengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.
Weakness:
I am pleased to see the authors' feedback on my previous questions and suggestions, and I believe the additional data analysis they have added is helpful. Here are my reserved concerns and newly discovered issues.
(1) Regarding the Inverted U-Shaped Curve In the revised manuscript, the authors have accepted some of my suggestions and conducted further analysis, which is now presented in Figure 3B. These results provide partial support for the authors' hypothesis. However, I still believe that the data from this study hardly convincingly support an inverted U-shaped distribution relationship.
The authors stated in their response, "it is challenging to determine the optimal level of ATL GABA," but I think this is achievable. From Figures 4C and 4D, the ATL GABA levels corresponding to the peak of the inverted U-shaped curve fall between 85 and 90. In my understanding, this can be considered as the optimal level of ATL GABA estimated based on the existing data and the inverted U-shaped curve relationship. However, in the latter half of the inverted U-shaped curve, there are quite few data points, and such a small number of data points hardly provides reliable support for the quantitative relationship in the latter half of the curve. I suggest that the authors should at least explicitly acknowledge this and be cautious in drawing conclusions. I also suggest that the authors consider fitting the data with more types of non-linear relationships, such as a ceiling effect (a combination of a slope and a horizontal line), or a logarithmic curve.(2) In Figure 2F, the authors demonstrated a strong practice effect in this study, which to some extent offsets the decrease in behavioral performance caused by cTBS. Therefore, I recommend that the authors give sufficient consideration to the practice effect in the data analysis.
One issue is the impact of the practice effect on the classification of responders and non-responders. Currently, most participants are classified as non-responders, suggesting that the majority of the population may not respond to the cTBS used in this study. This greatly challenges the generalizability of the experimental conclusions. However, the emergence of so many non-responders is likely due to the prominent practice effect, which offsets part of the experimental effect. If the practice effect is excluded, the number of responders may increase. The authors might estimate the practice effect based on the vertex simulation condition and reclassify participants after excluding the influence of the practice effect.
Another issue is that considering the significant practice effect, the analysis in Figure 4D, which mixes pre- and post-test data, may not be reliable.(3) The analysis in Figure 3A has a double dipping issue. Suppose we generate 100 pairs of random numbers as pre- and post-test scores, and then group the data based on whether the scores decrease or increase; the pre-test scores of the group with decreased scores will have a very high probability of being higher than those of the group with increased scores. Therefore, the findings in Figure 3A seem to be meaningless.
(4) The authors use IE as a behavioral measure in some analyses and use accuracy in others. I recommend that the authors adopt a consistent behavioral measure.
-
Reviewer #2 (Public review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data with their previous MRS study suggest an inverted …
Reviewer #2 (Public review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data with their previous MRS study suggest an inverted u-shape between GABA concentration and behavioral performance. These results help to better understand the neuromodulatory effects of non-invasive brain stimulation on task performance.
Strengths:
Multimodal assessment of neurostimulation effects on the behavioral, neurochemical, and neural levels. In particular, the link between GABA modulation and behavior is timely and potentially interesting.
Weaknesses:
The analyses are not sound. Some of the effects are very weak and not all conclusions are supported by the data since some of the comparisons are not justified. There is some redundancy with a previous paper by the same authors, so the novelty and contribution to the field are overall limited. A network approach might help here.
-
Reviewer #3 (Public review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on …
Reviewer #3 (Public review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on the semantic task. This is in line with their second prediction. Finally, the authors conducted additional analyses to explore the mechanistic link between ATL inhibitory GABAergic action and semantic task performance. They show that this link is best captured by an inverted U-shaped function as a result of a quadratic linear regression model. Fitting this model to their data indicates that increasing GABA levels led to better task performance as long as they were not excessively low or excessively high. This was first tested as a relationship between GABA levels in the ATL and semantic task performance; then the same analyses were performed on the pre and post-cTBS TMS stimulation data, showing the same pattern. These results are in line with the conclusions of the authors.
Comments on revisions:
The authors have comprehensively addressed my comments from the first round of review, and I consider most of their answers and the steps they have taken satisfactorily. Their insights prompted me to reflect further on my own knowledge and thinking regarding the ATL function.
I do, however, have an additional and hopefully constructive comment regarding the point made about the study focusing on the left instead of bilateral ATL. I appreciate the methodological complexities and the pragmatic reasons underlying this decision. Nevertheless, briefly incorporating the justification for this decision into the manuscript would have been beneficial for clarity and completeness. The presented argument follows an interesting logic; however, despite strong previous evidence supporting it, the approach remains based on an assumption. Given that the authors now provide the group-level fMRI results captured more comprehensively in Supplementary Figure 2, where the bilateral pattern of fMRI activation can be observed in the current data, the authors could have strengthened their argument by asserting that the activation related to the given semantic association task in this data was bilateral. This would imply that the TMS effects and associated changes in GABA should be similar for both sites. Furthermore, it is worth noting the approach taken by Pobric et al. (2007, PNAS), who stimulated a site located 10 mm posterior to the tip of the left temporal pole along the middle temporal gyrus (MTG) and not the bilateral ATL.
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This study focuses on the role of GABA in semantic memory and its neuroplasticity. The researchers stimulated the left ATL and control site (vertex) using cTBS, measured changes in GABA before and after stimulation using MRS, and measured changes in BOLD signals during semantic and control tasks using fMRI. They analyzed the effects of stimulation on GABA, BOLD, and behavioral data, as well as the correlation between GABA changes and BOLD changes caused by the stimulation. The authors also analyzed the relationship between individual differences in GABA levels and behavioral performance in the semantic task. They found that cTBS stimulation led to increased GABA levels and decreased BOLD activity in the …
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This study focuses on the role of GABA in semantic memory and its neuroplasticity. The researchers stimulated the left ATL and control site (vertex) using cTBS, measured changes in GABA before and after stimulation using MRS, and measured changes in BOLD signals during semantic and control tasks using fMRI. They analyzed the effects of stimulation on GABA, BOLD, and behavioral data, as well as the correlation between GABA changes and BOLD changes caused by the stimulation. The authors also analyzed the relationship between individual differences in GABA levels and behavioral performance in the semantic task. They found that cTBS stimulation led to increased GABA levels and decreased BOLD activity in the ATL, and these two changes were highly correlated. However, cTBS stimulation did not significantly change participants' behavioral performance on the semantic task, although behavioral changes in the control task were found after stimulation. Individual levels of GABA were significantly correlated with individuals' accuracy on the semantic task, and the inverted U-shaped (quadratic) function provides a better fit than the linear relationship. The authors argued that the results support the view that GABAergic inhibition can sharpen activated distributed semantic representations. They also claimed that the results revealed, for the first time, a non-linear, inverted-U-shape relationship between GABA levels in the ATL and semantic function, by explaining individual differences in semantic task performance and cTBS responsiveness
Strengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.
We appreciated R1’s positive evaluation of our manuscript.
Weaknesses:
Regarding the behavioral effects of GABA on semantic tasks, especially its impact on neuroplasticity, the results presented in the article are inadequate to support the claims made by the authors. There are three aspects of results related to this: 1) the effects of cTBS stimulation on behavior, 2) the positive correlation between GABA levels and semantic task accuracy, and 3) the nonlinear relationship between GABA levels and semantic task accuracy. Among these three pieces of evidence, the clearest one is the positive correlation between GABA levels and semantic task accuracy. However, it is important to note that this correlation already exists before the stimulation, and there are no results supporting that it can be modulated by the stimulation. In fact, cTBS significantly increases GABA levels but does not significantly improve performance on semantic tasks. According to the authors' interpretation of the results in Table 1, cTBS stimulation may have masked the practice effects that were supposed to occur. In other words, the stimulation decreased rather than enhanced participants' behavioral performance on the semantic task.
The stimulation effect on behavioral performance could potentially be explained by the nonlinear relationship between GABA and performance on semantic tasks proposed by the authors. However, the current results are also insufficient to support the authors' hypothesis of an inverted U-shaped curve. Firstly, in Figure 3C and Figure 3D, the last one-third of the inverted U-shaped curve does not have any data points. In other words, as the GABA level increases the accuracy of the behavior first rises and then remains at a high level. This pattern of results may be due to the ceiling effect of the behavioral task's accuracy, rather than an inverted U-shaped ATL GABA function in semantic memory. Second, the article does not provide sufficient evidence to support the existence of an optimal level of GABA in the ATL. Fortunately, this can be tested with additional data analysis. The authors can estimate, based on pre-stimulus data from individuals, the optimal level of GABA for semantic functioning. They can then examine two expectations: first, participants with pre-stimulus GABA levels below the optimal level should show improved behavioral performance after stimulation-induced GABA elevation; second, participants with pre-stimulus GABA levels above the optimal level should exhibit a decline in behavioral performance after stimulation-induced GABA elevation. Alternatively, the authors can categorize participants into groups based on whether their behavioral performance improves or declines after stimulation, and compare the pre- and post-stimulus GABA levels between the two groups. If the improvement group shows significantly lower pre-stimulus GABA levels compared to the decline group, and both groups exhibit an increase in GABA levels after stimulation, this would also provide some support for the authors' hypothesis.
Another issue in this study is the confounding of simulation effects and practice effects. According to the results, there is a significant improvement in performance after the simulation, at least in the control task, which the authors suggest may reflect a practice effect. The authors argue that the results in Table 1 suggest a similar practice effect in the semantic task, but it is masked by the simulation of the ATL. However, since no significant effects were found in the ANOVA analysis of the semantic task, it is actually difficult to draw a conclusion. This potential confound increases the risk in data analysis and interpretation. Specifically, for Figure 3D, if practice effects are taken into account, the data before and after the simulation should not be analyzed together.
We thank for the R1’s thoughtful comments. Due to the limited dataset, it is challenging to determine the optimal level of ATL GABA. Here, we re-grouped the participants into the responders and non-responders to address the issues R1 raised. It is important to note that we applied cTBS over the ATL, an inhibitory protocol, which decreases cortical excitability within the target region and semantic task performance (Chiou et al., 2014; Jung and Lambon Ralph, 2016). Therefore, responders and non-responders were classified according to their semantic performance changes after the ATL stimulation: subjects showing a decrease in task performance at the post ATL cTBS compared to the baseline were defined as responders; whereas subjects showing no changes or an increase in their task performance after the ATL cTBS were defined as non-responders. Here, we used the inverse efficiency (IE) score (RT/1-the proportion of errors) as individual semantic task performance to combine accuracy and RT. Accordingly, we had 7 responders and 10 non-responders.
Recently, we demonstrated that the pre-stimulation neurochemical profile of the ATL was associated with cTBS responsiveness on semantic processing (Jung et al., 2022). Specifically, the baseline GABA and Glx levels in the ATL predicted cTBS induced semantic task performance changes: individuals with higher GABA and lower Glx in the ATL would show bigger inhibitory effects and responders who decreased semantic task performance after ATL stimulation. Importantly, the baseline semantic task performance was significantly better in responders compared to non-responders. Thus, we expected that responders would show better semantic task performance along with higher ATL GABA levels in their pre-stimulation session relative to non-responders. We performed the planned t-tests to examine the difference in task performance and ATL GABA levels in pre-stimulation session. The results revealed that responders had lower IE (better task performance, t = -1.756, p = 0.050) and higher ATL GABA levels (t = 2.779, p = 0.006) in the pre-stimulation session (Figure 3).
In addition, we performed planned paired t-test to investigate the cTBS effects on semantic task performance and regional ATL GABA levels according to the groups (responders and non-responders). Responders showed significant increase of IE (poorer performance, t = -1.937, p = 0.050) and ATL GABA levels (t = -2.203, p = 0.035) after ATL cTBS. Non-responders showed decreased IE (better performance, t = 2.872, p = 0.009) and increased GABA levels in the ATL (t = -3.912, p = 0.001) after the ATL stimulation. The results were summarised in Figure 3.
It should be noted that there was no difference between the responders and non-responders in the control task performance at the pre-stimulation session. Both groups showed better performance after the ATL stimulation – practice effects (Author response image 1 below).
Author response image 1.
As we expected, our results replicated the previous findings (Jung et al., 2022) that responders who showed the inhibitory effects on semantic task performance after the ATL stimulation had higher GABA levels in the ATL than non-responders at their baseline, the pre-stimulation session. Importantly, cTBS increased ATL GABA levels in both responders and non-responders. These findings support our hypothesis – the inverted U-shaped ATL GABA function for cTBS response (Figure 4B). cTBS over the ATL resulted in the inhibition of semantic task performance among individuals initially characterized by higher concentrations of GABA in the ATL, indicative of better baseline semantic capacity. Conversely, the impact of cTBS on individuals with lower semantic ability and relatively lower GABA levels in the ATL was either negligible or exhibited a facilitatory effect. This study posits that individuals with elevated GABA levels in the ATL tend to be more responsive to cTBS, displaying inhibitory effects on semantic task performance (responders). On the contrary, those with lower GABA concentrations and reduced semantic ability were less likely to respond or even demonstrated facilitatory effects following ATL cTBS (non-responders). Moreover, our findings suggest the critical role of the baseline neurochemical profile in individual responsiveness to cTBS in the context of semantic memory. This highlights substantial variability among individuals in terms of semantic memory and its plasticity induced by cTBS.
Our analyses with responders and non-responders have highlighted significant inter-individual variability in both pre- and post-ATL stimulation sessions, including behavioural outcomes and ATL GABA levels. Responders showed distinctive neurochemical profiles in the ATL, associating with their task performance and responsiveness to cTBS in semantic memory. Our findings suggest that responders may possess an optimal level of ATL GABA conducive to efficient semantic processing. This results in enhanced semantic task performance and increased responsiveness to cTBS, leading to inhibitory effects on semantic processing following an inverted U-shaped function. On the contrary, non-responders, characterized by relatively lower ATL GABA levels, exhibited poorer semantic task performance compared to responders at the baseline. The cTBS-induced increase in GABA may contribute to their subsequent improvement in semantic performance. These results substantiate our hypothesis regarding the inverted U-shape function of ATL GABA and its relationship with semantic behaviour.
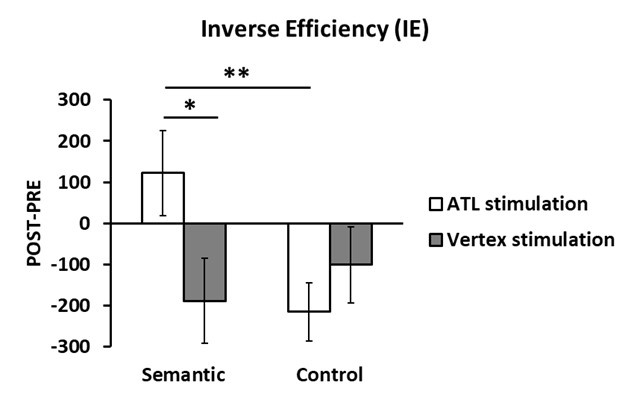
To address the confounding of simulation effects and practice effects in behavioural data, we used the IE and computed cTBS-induced performance changes (POST-PRE). Employing a 2 x 2 ANOVA with stimulation (ATL vs. Vertex) and task (Semantic vs. Control) as within subject factors, we found a significant task effect (F1, 15 = 6.656, p = 0.021) and a marginally significant interaction between stimulation and task (F1, 15 = 4.064, p = 0.061). Post hoc paired t-test demonstrated that ATL stimulation significantly decreased semantic task performance (positive IE) compared to both vertex stimulation (t = 1.905, p = 0.038) and control task (t = 2.814, p = 0.006). Facilitatory effects (negative IE) were observed in the control stimulation and control task. Please, see the Author response image 2 below. Thus, we believe that ATL cTBS induced task-specific inhibitory effects in semantic processing.
Author response image 2.
Accordingly, we have revised the Methods and Materials (p 25, line 589), Results (p8, line 188, p9-11, line 202- 248), Discussion (p19, line 441) and Figures (Fig. 2-3 & all Supplementary Figures).
Reviewer #2 (Public Review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data from their previous MRS study suggest an inverted U-shape between GABA concentration and behavioral performance. These results help to better understand the neuromodulatory effects of non-invasive brain stimulation on task performance.
Strengths:
Multimodal assessment of neurostimulation effects on the behavioral, neurochemical, and neural levels. In particular, the link between GABA modulation and behavior is timely and potentially interesting.
We appreciated R2’s positive evaluation of our manuscript.
Weaknesses:
The analyses are not sound. Some of the effects are very weak and not all conclusions are supported by the data since some of the comparisons are not justified. There is some redundancy with a previous paper by the same authors, so the novelty and contribution to the field are overall limited. A network approach might help here.
Thank you for your thoughtful critique. We have taken your comments into careful consideration and have made efforts to address them.
We acknowledge the limitations regarding the strength of some effects and the potential lack of justification for certain conclusions drawn from the data. In response, we have reviewed our analyses and performed new analyses to address the behavioural discrepancies and strengthened the justifications for our conclusions.
Regarding the redundancy with a previous paper by the same authors, we understand your concern about the novelty and contribution to the field. We aim to clarify the unique contributions of our current study compared to our previous work. The main novelty lies in uncovering the neurochemical mechanisms behind cTBS-induced neuroplasticity in semantic representation and establishing a non-linear relationship between ATL GABA levels and semantic representation. Our previous work primarily demonstrated the linear relationship between ATL GABA levels and semantic processing. In the current study, we aimed to address two key objectives: 1) investigate the role of GABA in the ATL in short-term neuroplasticity in semantic representation, and 2) explore a biologically more plausible function between ATL GABA levels and semantic function using a larger sample size by combining data from two studies.
Additionally, we appreciate your suggestion regarding a network approach. We have explored the relationship between ATL GABA and cTBS-induced functional connectivity changes in our new analysis. However, there was no significant relationship between them. In the current study, our decision to focus on the mechanistic link between ATL GABA, task-induced activity, and individual semantic task performance reflects our intention to provide a detailed exploration of the role of GABA in the ATL and semantic neuroplasticity.
We have addressed the specific weaknesses raised by Reviewer #2 in detail in our response to 'Reviewer #2 Recommendations For The Authors'.
Reviewer #3 (Public Review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on the semantic task. This is in line with their second prediction. Finally, the authors conducted additional analyses to explore the mechanistic link between ATL inhibitory GABAergic action and semantic task performance. They show that this link is best captured by an inverted U-shaped function as a result of a quadratic linear regression model. Fitting this model to their data indicates that increasing GABA levels led to better task performance as long as they were not excessively low or excessively high. This was first tested as a relationship between GABA levels in the ATL and semantic task performance; then the same analyses were performed on the pre and post-cTBS TMS stimulation data, showing the same pattern. These results are in line with the conclusions of the authors.
Strengths:
I thoroughly enjoyed reading the manuscript and appreciate its contribution to the field of the role of the ATL in semantic processing, especially given the efforts to overcome the immense challenges of investigating ATL function by neuroscientific methods such as MRS, fMRI & TMS. The main strengths are summarised as follows:
• The work is methodologically rigorous and dwells on complex and complementary multimethod approaches implemented to inform about ATL function in semantic memory as reflected in changes in regional GABA concentrations. Although the authors previously demonstrated a negative relationship between increased GABA levels and BOLD signal changes during semantic processing, the unique contribution of this work lies within evidence on the effects of cTBS TMS over the ATL given by direct observations of GABA concentration changes and further exploring inter-individual variability in ATL neuroplasticity and consequent semantic task performance.
• Another major asset of the present study is implementing a quadratic regression model to provide insights into the non-linear relationship between inhibitory GABAergic activity within the ATLs and semantic cognition, which improves with increasing GABA levels but only as long as GABA levels are not extremely high or low. Based on this finding, the authors further pinpoint the role of inter-individual differences in GABA levels and cTBS TMS responsiveness, which is a novel explanation not previously considered (according to my best knowledge) in research investigating the effect of TMS on ATLs.
• There are also many examples of good research practice throughout the manuscript, such as the explicitly stated exploratory analyses, calculation of TMS electric fields, using ATL optimised dual echo fRMI, links to open source resources, and a part of data replicates a previous study by Jung et. al (2017).
We appreciated R3’s very positive evaluation of our manuscript.
Weaknesses:
• Research on the role of neurotransmitters in semantic memory is still very rare and therefore the manuscript would benefit from more context on how GABA contributes to individual differences in cognition/behaviour and more justification on why the focus is on semantic memory. A recommendation to the authors is to highlight and explain in more depth the particular gaps in evidence in this regard.
This is an excellent suggestion. Accordingly, we have revised our introduction, highlighting the role of GABA on individual differences in cognition and behaviour and research gap in this field.
Introduction p3, line 77
“Research has revealed a link between variability in the levels of GABA in the human brain and individual differences in cognitive behaviour (for a review, see 5). Specifically, GABA levels in the sensorimotor cortex were found to predict individual performance in the related tasks: higher GABA levels were correlated with a slower reaction time in simple motor tasks (12) as well as improved motor control (13) and sensory discrimination (14, 15). Visual cortex GABA concentrations were positively correlated with a stronger orientation illusion (16), a prolonged binocular rivalry (17), while displaying a negative correlation with motion suppression (17). Individuals with greater frontal GABA concentrations demonstrated enhanced working memory capacity (18, 19). Studies on learning have reported the importance of GABAergic changes in the motor cortex for motor and perceptual learning: individuals showing bigger decreases in local GABA concentration can facilitate this plasticity more effectively (12, 20-22). However, the relationship between GABAergic inhibition and higher cognition in humans remains unclear. The aim of the study was to investigate the role of GABA in relation to human higher cognition – semantic memory and its neuroplasticity at individual level.”
• The focus across the experiments is on the left ATL; how do the authors justify this decision? Highlighting the justification for this methodological decision will be important, especially given that a substantial body of evidence suggests that the ATL should be involved in semantics bilaterally (e.g. Hoffman & Lambon Ralph, 2018; Lambon Ralph et al., 2009; Rice et al., 2017; Rice, Hoffman, et al., 2015; Rice, Ralph, et al., 2015; Visser et al., 2010).
This is an important point, which we thank R3 for. Supporting the bilateral ATL systems in semantic representation, previous rTMS studies delivered an inhibitory rTMS in the left and right ATL and both ATL stimulation significantly decreased semantic task performance (Pobric et al., 2007 PNAS; 2010 Neuropsychologia; Lambon Ralph et al., 2009 Cerebral Cortex). Importantly, there was no significant difference on rTMS effects between the left and right ATL stimulation. Therefore, we assume that either left or right ATL stimulation could produce similar, intended rTMS effects on semantic processing. In the current study, we combined the cTBS with multimodal imaging to examine the cTBS effects in the ATL. Due to the design of the study (having a control site, control task, and control stimulation) and limitation of scanning time, we could have a target region for the simulation and chose the left ATL, which was the same MRS VOI of our precious study (Jung et al., 2017). This enabled us to combine the datasets to explore GABAergic function in the ATL.
• When describing the results, (Pg. 11; lines 233-243), the authors first show that the higher the BOLD signal intensity in ATL as a response to the semantic task, the lower the GABA concentration. Then, they state that individuals with higher GABA concentrations in the ATL perform the semantic task better. Although it becomes clearer with the exploratory analysis described later, at this point, the results seem rather contradictory and make the reader question the following: if increased GABA leads to less task-induced ATL activation, why at this point increased GABA also leads to facilitating and not inhibiting semantic task performance? It would be beneficial to acknowledge this contradiction and explain how the following analyses will address this discrepancy.
We apologised that our description was not clear. As R1 also commented this issue, we re-analysed behavioural results and demonstrated inter-individual variability in response to cTBS (Please, see the reply to R1 above).
• There is an inconsistency in reporting behavioural outcomes from the performance on the semantic task. While experiment 1 (cTBS modulates regional GANA concentrations and task-related BOLD signal changes in the ATL) reports the effects of cTBS TMS on response times, experiment 2 (Regional GABA concentrations in the ATL play a crucial role in semantic memory) and experiment 3 (The inverted U-shaped function of ATL GABA concentration in semantic processing) report results on accuracy. For full transparency, the manuscript would benefit from reporting all results (either in the main text or supplementary materials) and providing further explanations on why only one or the other outcome is sensitive to the experimental manipulations across the three experiments.
Regarding the inconsistency of behavioural outcome, first, there were inter- individual differences in our behavioural data (see the Figure below). Our new analyses revealed that there were responders and non-responders in terms of cTBS responsiveness (please, see the reply to R1 above. It should be noted that the classification of responders and non-responders was identical when we used semantic task accuracy). In addition, RT was compounded by practice effects (faster in the post-stimulation sessions), except for the ATL-post session. Second, we only found the significant relationship between semantic task accuracy and ATL GABA concentrations in both previous (Jung et al., 2017) and current study. ATL GABA levels were not correlated with semantic RT (Jung et al., 2017: r = 0.34, p = 0.14, current study: r = 0.26, p = 0.14). It should be noted that there were no significant correlations between ATL GABA levels and semantic inverse efficiency (IE) in both studies (Jung et al., 2017: r = 0.13, p = 0.62, current study: r = 0.22, p = 0.44). As a result, we found no significant linear and non-linear relationship between ATL GABA levels and RT (linear function R2 = 0.21, p =0.45, quadratic function: R2 = 0.17, p = 0.21) and between ATL GABA levels and IE (linear function R2 = 0.24, p =0.07, quadratic function: R2 = 2.24, p = 0.12). Thus, our data suggests that GABAergic action in the ATL may sharpen activated distributed semantic representations through lateral inhibition, leading to more accurate semantic performance (Isaacson & Scanziani., 2011; Jung et al., 2017).
We agreed with R3’s suggestion to report all results. The results of control task and control stimulation were included in Supplementary information (Figure S1, S4-5).
Overall, the most notable impact of this work is the contribution to a better understanding of individual differences in semantic behaviour and the potential to guide therapeutic interventions to restore semantic abilities in neurological populations. While I appreciate that this is certainly the case, I would be curious to read more about how this could be achieved.
Thank you once again to R3 for the positive evaluation of our study. We acknowledge your interest in understanding the practical implications of our findings. It is crucial to highlight the substantial variability in the effectiveness of rTMS and TBS protocols among individuals. Previous studies in healthy subjects have reported response rates ranging from 40% to 70% in the motor cortex, and in patients, the remission rate for rTMS treatment in treatment-resistant depression is around 29%. Presently, the common practice in rTMS treatment is to apply the same protocol uniformly to all patients.
Our study demonstrated that 40% of individuals in our sample were classified as responders to ATL cTBS. Notably, we observed differences in ATL GABA levels before stimulation between responders and non-responders. Responders exhibited higher baseline ATL GABA levels, along with better semantic performance at the baseline (as mentioned in our response to R1). This suggests that establishing the optimal level of ATL GABA by assessing baseline GABA levels before stimulation could enable the tailoring of an ideal protocol for each individual, thereby enhancing their semantic capability. To achieve this, more data is needed to delineate the proposed inverted U-shaped function of ATL GABA in semantic memory.
Our ongoing efforts involve collecting additional data from both healthy aging and dementia cohorts using the same protocol. Additionally, future pharmacological studies aim to modulate GABA, providing a deeper understanding of the individual variations in semantic function. These initiatives contribute to the potential development of personalized therapeutic interventions for individuals with semantic impairments.
Reviewer #1 (Recommendations For The Authors):
My major suggestion is to include an analysis regarding the "existence of an optimal GABA level". This would be the most direct test for the authors' hypothesis on the relationship between GABA and semantic memory and its neuroplasticity. Please refer to the public review section for details.
Here are some other suggestions and questions.
(1) The sample size of this study is relatively small. Although the sample size was estimated, a small sample size can bring risks to the generalizability of the results to the population. How did the author consider this risk? Is it necessary to increase the sample size?
We agreed with R1’s comments. However, the average of sample size in healthy individuals was 17.5 in TMS studies on language function (number of studies = 26, for a review, see Qu et al, 2022 Frontiers in Human Neuroscience), 18.3 in the studies employing rTMS and fMRI on language domain (number of studies = 8, for a review, see Hartwigsen & Volz., 2021 NeuroImage), and 20.8 in TMS combined MRS studies (number of studies = 11, for a review, see Cuypers & Marsman., 2021 NeuroImage). Notably, only two studies utilizing rTMS, fMRI, and MRS had sample sizes of N = 7 (Grohn et al., 2019 Frontiers in Neuroscience) and N = 16 (Rafique & Steeves. 2020 Brain and Behavior). Despite having 19 participants in our current study, it is noteworthy that our sample size aligns closely with studies employing similar approaches and surpasses those employing the same methodology.
As a result of the changes in a scanner and the relocation of the authors to different institutes, it is impossible to increase the sample size for this study.
(2) How did the authors control practice effects? How many practice trials were arranged before the experiment? Did you avoid the repetition of stimuli in tasks before and after the stimuli?
At the beginning of the experiment, participants performed the practice session (20 trials) for each tasks outside of the scanner. Stimuli in tasks were not repeated before and after stimulation sessions.
(3) In Figures 2D and E, does the vertical axis of the BOLD signal refer to the semantic task itself or the difference between the semantic and control tasks? Could you provide the respective patterns of the BOLD signal before and after the stimuli in the semantic and control tasks in a figure?
We apologised that the names of axis of Figure 2 were not clear. In Fig 2D-E, the BOLD signal changes refer to the semantic task itself. Accordingly, we have revised the Fig. 2.
(4) Figure 1A shows that MRS ATL always comes before MRS Vertex. Was the order of them counterbalanced across participants?
The order of MRS acquisition was not counterbalanced across participants.
(5) I am confused by the statement "Our results provide strong evidence that regional GABA levels increase following inhibitory cTBS in the human associative cortex, specifically in the ATL, a representational semantic hub. Notably, the observed increase was specific to the ATL and semantic processing, as it was not observed in the control region (vertex) and not associated with control processing (visuospatial processing)". GABA levels are obtained in the MRS, and this stage does not involve any behavioral tasks. Why do the authors state that the increase in GABA levels was specific to semantic processing and was not associated with control processing?
Following R1’s suggestion, we have re-analysed behavioural data and showed cTBS-induced suppression in semantic task performance after ATL stimulation only (please, see the reply above). There were no cTBS effects in the control task performance, control site (vertex) and no correlations between the ATL GABA levels and control task performance. The Table was added to the Supplementary Information as Table S3.
(6) In Figure 3, the relationship between GABA levels in the ATL and performance on semantic tasks is presented. What is the relationship between GABA levels at the control site and performance on semantic tasks? Should a graph be provided to illustrate this?
As the vertex was not involved in semantic processing (no activation during semantic processing), we did not perform the analysis between vertex GABA levels and semantic task performance. Following R3’s suggestion, we performed a linear regression between vertex GABA levels and semantic task performance in the pre-stimulation session, accounting for GM volume, age, and sex. As we expected that there was no significant relationship between them. (R2 = 0.279, p = 0.962).
(7) The author claims that GABA can sharpen distributed semantic representations. However, even though there is a positive correlation between GABA levels and semantic performance, there is no direct evidence supporting the inference that this correlation is achieved through sharpening distributed semantic representations. How did the author come to this conclusion? Are there any other possibilities?
We showed that ATL GABA concentrations in pre-stimulation was ‘negatively’ correlated with task-induced regional activity in the ATL and ‘positively’ correlated with semantic task performance. In our semantic task, such as recognizing a camel (Fig. 1), the activation of all related information in the semantic representation (e.g., mammal, desert, oasis, nomad, humps, & etc.) occurs. To respond accurately to the task (a cactus), it becomes essential to suppress irrelevant meanings through an inhibitory mechanism. Therefore, the inhibitory processing linked to ATL GABA levels may contribute to more efficient processing in this task.
Animal studies have proposed a related hypothesis in the context of the close interplay between activation and inhibition in sensorimotor cortices (Isaacson & Scanziani., 2011). Liu et al (2011, Neuron) demonstrated that the rise of excitatory glutamate in the visual cortex is followed by the increase of inhibitory GABA in response to visual stimuli. Tight coupling of these paired excitatory-inhibitory functions results in a sharpening of the activated representation. (for a review, see Isaacson & Scanziani., 2011 Neuron How Inhibition Shapes Cortical Activity). In human, Kolasinski et al (2017, Current Biology) revealed that higher sensorimotor GABA levels are associated with more selective cortical tuning measured fMRI, which in turn is associated with enhanced perception (better tactile discrimination). They claimed that the relationship between inhibition and cortical tuning could result from GABAergic signalling, shaping the selective response profiles of neurons in the primary sensory regions of the brain. This process is crucial for the topographic organization (task-induced fMRI activation in the sensorimotor cortex) vital to sensory perception.
Building on these findings, we suggest a similar mechanism may operate in higher-order association cortices, including the ATL semantic hub. This suggests a process that leads to more sharply defined semantic representations associated with more selective task-induced activation in the ATL and, consequently, more accurate semantic performance (Jung et al., 2017).
Reviewer #2 (Recommendations For The Authors):
Major issues:
(1) It wasn't completely clear what the novel aspect of this study relative to their previous one on GABAergic modulation in semantic memory issue, this should be clarified. If I understand correctly, the main difference from the previous study is that this study considers the TMS-induced modulation of GABA?
We apologise that the novelty of study was not clear. The main novelty lies in uncovering the neurochemical mechanisms behind cTBS-induced neuroplasticity in semantic representation and establishing a non-linear relationship between ATL GABA levels and semantic representation. Our previous work firstly demonstrated the linear relationship between the ATL GABA levels and semantic processing. In the current study, we aimed to address two key objectives: 1) investigate the role of GABA in the ATL in short-term neuroplasticity in semantic representation, and 2) explore a biologically more plausible function between ATL GABA levels and semantic function using a larger sample size by combining data from two studies.
The first part of the experiment in this study mirrored our previous work, involving multimodal imaging during the pre-stimulation session. We conducted the same analysis as in our previous study to replicate the findings in a different cohort. Subsequently, we combined the data from both studies to examine the potential inverted U-shape function between ATL GABA levels and semantic function/neuroplasticity.
Accordingly, we have revised the Introduction by adding the following sentences.
“The study aimed to investigate the neural mechanisms underlying cTBS-induced neuroplasticity in semantic memory by linking cortical neurochemical profiles, task-induced regional activity, and variability in semantic memory capability within the ATL.”
“Furthermore, to address and explore the relationship between regional GABA levels in the ATL and semantic memory function, we combined data from our previous study (Jung et al., 2017) with the current study’s data.”
(2) I found the scope of the study very narrow. I guess everyone agrees that TMS induces network effects, but the authors selectively focus on the modulation in the ATL. This is unfortunate since semantic memory requires the interaction between several brain regions and a network perspective might add some novel aspect to this study which has a strong overlap with their previous one. I am aware that MRS can only measure pre-defined voxels but even these changes could be related to stimulation-induced effects on task-related activity at the whole brain level.
We appreciate R2's thoughtful comments and acknowledge the concern about the perceived narrow scope of the study. We agreed with the notion that cTBS induces network-level changes. In our investigation, we did observe cTBS over the ATL influencing task-induced regional activity in other semantic regions and functional connectivity within the semantic system. Specifically, ATL cTBS increased activation in the right ATL after ATL stimulation compared to pre-stimulation, along with increased functional connectivity between the left and right ATL, between the left ATL and right semantic control regions (IFG and pMTG), and between the left ATL and right angular gyrus. These results were the replication of Jung & Lambon Ralph (2016) Cerebral Cortex.
However, it is important to note that we did not find any significant correlations between ATL GABA changes and cTBS-induced changes in the functional connectivity. Consequently, we are currently preparing another paper that specifically addresses the network-level changes induced by ATL cTBS. In the current study, our decision to focus on the mechanistic link between ATL GABA, task-induced activity, and individual semantic task performance reflects our intention to provide a detailed exploration of the role of GABA in the ATL and semantic neuroplasticity.
(3) On a related note, I think the provided link between GABAergic modulation and behavioral changes after TMS is somehow incomplete because it ignores the stimulation effects on task-related activity. Could these be linked in a regression analysis with two predictors (with behavior or GABA level as a criterion and the other two variables as predictors)?
In response to R2’s suggestion, we performed a multiple regression analysis, by modelling cTBS-induced ATL GABA changes (POST-PRE), task-related BODL signal changes (POST-PRE), and semantic task performance (IE) changes (POST-PRE). The model with GABA changes (POST-PRE) as a criterion was significant (F2, 14 = 8.77, p = 0.003), explaining 56% of cTBS-induced ATL GABA changes (adjusted R2) with cTBS-related ATL BOLD signal changes and semantic task performance changes. However, the model with semantic task performance change (POST-PRE) as a criterion was not significant (F = 0.26, p = 0.775). Therefore, cTBS-induced changes in ATL BOLD signals and semantic task performance significantly predicted the cTBS-induced ATL GABA changes. It was found that cTBS-induced ATL BOLD signal changes significantly predicted cTBS-induced GABA changes in the ATL (β = -4.184, p = 0.001) only, aligning with the results of our partial correlation analysis.
Author response table 1.

(4) Several statements in the intro and discussion need to be rephrased or toned down. For example, I would not agree that TBS "made healthy individuals mimic semantic dementia patients". This is clearly overstated. TMS protocols slightly modulate brain functions, but this is not similar to lesions or brain damage. Please rephrase. In the discussion, it is stated that the results provide "strong evidence". I disagree based on the overall low values for most comparisons.
Hence, we have revised both the Introduction and the Discussion.
“Perturbing the ATL with inhibitory repetitive transcranial magnetic stimulation (rTMS) and theta burst stimulation (TBS) resulted in healthy individuals exhibiting slower reaction times during semantic processing.”
“Our results demonstrated an increase in regional GABA levels following inhibitory cTBS in human associative cortex, specifically in the ATL, a representational semantic hub.”
(5) Changes in the BOLD signal in the ATL: There is a weak interaction between stimulation and VOI and post hoc comparisons with very low values reported. Are these corrected for multiple comparisons? I think that selectively reporting weak values with small-volume corrections (if they were performed) does not provide strong evidence. What about whole-brain effects and proper corrections for multiple comparisons?
There was no significant interaction between the stimulation (ATL vs. Vertex) and session (pre vs post) in the ATL BOLD signal changes (p = 0.29). Our previous work combining rTMS with fMRI (Binney et al., 2015; Jung & Lambon Ralph, 2016) demonstrated that there was no significant rTMS effects on the whole brain analysis and only ROI analyses revealed the subtle but significant rTMS effects in the target site (reduction of task-induced ATL activity). In the current study, we focused our hypothesis on the anticipated decrease in task-induced regional activity in the ATL during semantic processing following the inhibitory cTBS. Accordingly, we conducted planned paired t-tests specifically within the ATL for BOLD signal changes without applying multiple comparison corrections. It's noted that these results were derived from regions of interest (ROIs) and not from small-volume corrections. Furthermore, no significant findings emerged from the comparison of the ATL post-session vs. Vertex post-session and the ATL pre-session vs. ATL post-session in the whole-brain analysis (see Supplementary figure 2).
Accordingly, we have added the Figure S2 in the Supplementary Information.
(6) Differences between selected VOIs: Numerically, the activity (BOLD signal effect) is higher in the vertex than the ATL, even in the pre-TMS session (Figure 2D). What does that mean? Does that indicate that the vertex also plays a role in semantic memory?
We apologise that the figure was not clear. Fig. 2D displays the BOLD signal changes in the ATL VOI for the ATL and Vertex stimulation. As there was no activation in the vertex during semantic processing, we did not present the fMRI results of vertex VOI (please, see Author response image 3 below). Accordingly, we have revised the label of Y axis of the Figure 2D – ATL BOLD signal change.
Author response image 3.
The cTBS effects within the Vertex VOI during semantic processing
(7) Could you provide the e-field for the vertex condition?
We have added it in the Supplementary Information as Supplementary Figure 6.
(8) Stimulation effects on performance (RTs): There is a main effect of the session in the control task. Post-hoc tests show that control performance is faster in the post-pre comparison, while the semantic task is not faster after ATL TMS (as it might be delayed). I think you need to perform a 3-way ANOVA here including the factor task if you want to show task specificity (e.g., differences for the control but not semantic task) and then a step-down ANOVA or t-tests.
Thanks for R2’s suggestion. We have addressed this issue in reply to R1. Please, see the reply to R1 for semantic task performance analysis.
Minor issue:
In the visualization of the design, it would be helpful to have the timing/duration of the different measures to directly understand how long the experiment took.
We have added the duration of the experiment design in the Figure 1.
Reviewer #3 (Recommendations For The Authors):
Further Recommendations:
• Pg. 6; lines 138-147: There is a sense of uncertainty about the hypothesis conveyed by expressions such as 'may' or 'could be'. A more confident tone would be beneficial.
Thanks for R3’s thoughtful suggestion. We have revised the Introduction.
• Pg. 6; line 155: left or bilateral ATL, please specify.
We have added ‘left’ in the manuscript.
• Pg. 8; line 188: Can the authors provide a table with peak activations to complement the figure?
We have added the Table for the fMRI results in the Supplementary Information (Table S1).
• Pg 9; Figure 2C: The ATL activation elicited by the semantic task seems rather medial. What are the exact peak coordinates for this cluster, and how can the authors demonstrate that the electric fields induced by TMS, which seem rather lateral (Figure 2A), also impacted this area? Please explain.
We apologise that the Figure was not clear. cTBS was delivered to the peak coordinate of the left ventral ATL [-36, -15, -30] determined by previous fMRI studies (Binney et al., 2010; Visser et al., 2012). To confirm the cTBS effects at the target region, we conducted ROI analysis centred in the ventral ATL [-36, -15, -30] and the results demonstrated a reduced ATL activity after ATL stimulation during semantic processing (t = -2.43, p = 0.014) (please, see Author response image 4 below). Thus, cTBS successfully modulated the ATL activity reaching to the targe coordinate.
Author response image 4.
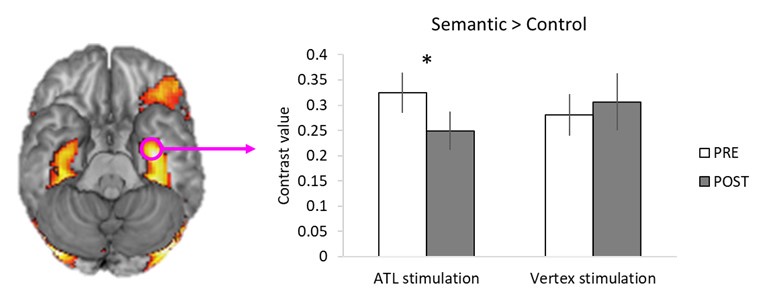
• Pg.23; line 547: What was the centre coordinate of the ROI (VOI), and was it consistent across all participants? Please specify.
We used the ATL MRS VOI (a hexahedron with 4cm x 2cm x 2cm) for our regions of interest analysis and the central coordinate was around -45, -12, -20 (see Author response image 5). As we showed in Fig. 1C, the location of ATL VOI was consistent across all participants.
Author response image 5.
• Pg. 24; line 556-570: What software was used for performing the statistical analyses? Please specify.
We have added the following sentence.
“Statistical analyses were undertaken using Statistics Package for the Social Sciences (SPSS, Version 25, IBM Cary, NC, USA) and RStudio (2023).”
• Pg. 21; line 472-480: It is not clear if and how neuronavigation was used (e.g. were T1scans or an average MNI template used, what was the exact coordinate of stimulation and how was it decided upon). Please specify.
We apologised the description was not clear. We have added a paragraph describing the procedure.
“The target site in the left ATL was delineated based on the peak coordinate (MNI -36 -15 -30), which represents maximal peak activation observed during semantic processing in previous distortion-corrected fMRI studies (38, 41). This coordinate was transformed to each individual’s native space using Statistical Parametric Mapping software (SPM8, Wellcome Trust Centre for Neuroimaging, London, UK). T1 images were normalised to the MNI template and then the resulting transformations were inverted to convert the target MNI coordinate back to the individual's untransformed native space coordinate. These native-space ATL coordinates were subsequently utilized for frameless stereotaxy, employing the Brainsight TMS-MRI co-registration system (Rogue Research, Montreal, Canada). The vertex (Cz) was designated as a control site following the international 10–20 system.”
• Miscellaneous
- line 57: insert 'about' to the following sentence: '....little is known the mechanisms linking'
- line 329: 'Previous, we demonstrated'....should be Previously we demonstrated....
We thank for R3’s thorough evaluation our manuscript. We have revised them.
Furthermore, it would be an advantage to make the data freely available for the benefit of the broader scientific community.
We appreciate Reviewer 3’s suggestion. Currently, this data is being used in other unpublished work. However, upon acceptance of this manuscript, we will make the data freely available for the benefit of the broader scientific community.
Chiou R, Sowman PF, Etchell AC, Rich AN (2014) A conceptual lemon: theta burst stimulation to the left anterior temporal lobe untangles object representation and its canonical color. J Cogn Neurosci 26:1066-1074.
Jung J, Lambon Ralph MA (2016) Mapping the Dynamic Network Interactions Underpinning Cognition: A cTBS-fMRI Study of the Flexible Adaptive Neural System for Semantics. Cereb Cortex 26:3580-3590.
Jung J, Williams SR, Sanaei Nezhad F, Lambon Ralph MA (2017) GABA concentrations in the anterior temporal lobe predict human semantic processing. Sci Rep 7:15748.
Jung J, Williams SR, Nezhad FS, Lambon Ralph MA (2022) Neurochemical profiles of the anterior temporal lobe predict response of repetitive transcranial magnetic stimulation on semantic processing. Neuroimage 258:119386.
-
-
-
eLife assessment
This is a valuable paper that might contribute new insight into the role of GABA in semantic memory, which is a significant question in higher cognition. However, the empirical support for the main claims is incomplete, with some results not fully coherent and robust – the paper would benefit from more rigorous analyses. These results, once strengthened, will be of interest to broad readers of the neuroscience and cognitive neuroscience community.
-
Reviewer #1 (Public Review):
Summary:
This study focuses on the role of GABA in semantic memory and its neuroplasticity. The researchers stimulated the left ATL and control site (vertex) using cTBS, measured changes in GABA before and after stimulation using MRS, and measured changes in BOLD signals during semantic and control tasks using fMRI. They analyzed the effects of stimulation on GABA, BOLD, and behavioral data, as well as the correlation between GABA changes and BOLD changes caused by the stimulation. The authors also analyzed the relationship between individual differences in GABA levels and behavioral performance in the semantic task. They found that cTBS stimulation led to increased GABA levels and decreased BOLD activity in the ATL, and these two changes were highly correlated. However, cTBS stimulation did not …Reviewer #1 (Public Review):
Summary:
This study focuses on the role of GABA in semantic memory and its neuroplasticity. The researchers stimulated the left ATL and control site (vertex) using cTBS, measured changes in GABA before and after stimulation using MRS, and measured changes in BOLD signals during semantic and control tasks using fMRI. They analyzed the effects of stimulation on GABA, BOLD, and behavioral data, as well as the correlation between GABA changes and BOLD changes caused by the stimulation. The authors also analyzed the relationship between individual differences in GABA levels and behavioral performance in the semantic task. They found that cTBS stimulation led to increased GABA levels and decreased BOLD activity in the ATL, and these two changes were highly correlated. However, cTBS stimulation did not significantly change participants' behavioral performance on the semantic task, although behavioral changes in the control task were found after stimulation. Individual levels of GABA were significantly correlated with individuals' accuracy on the semantic task, and the inverted U-shaped (quadratic) function provides a better fit than the linear relationship. The authors argued that the results support the view that GABAergic inhibition can sharpen activated distributed semantic representations. They also claimed that the results revealed, for the first time, a non-linear, inverted-U-shape relationship between GABA levels in the ATL and semantic function, by explaining individual differences in semantic task performance and cTBS responsivenessStrengths:
The findings of the research regarding the increase of GABA and decrease of BOLD caused by cTBS, as well as the correlation between the two, appear to be reliable. This should be valuable for understanding the biological effects of cTBS.Weaknesses:
Regarding the behavioral effects of GABA on semantic tasks, especially its impact on neuroplasticity, the results presented in the article are inadequate to support the claims made by the authors. There are three aspects of results related to this: 1) the effects of cTBS stimulation on behavior, 2) the positive correlation between GABA levels and semantic task accuracy, and 3) the nonlinear relationship between GABA levels and semantic task accuracy. Among these three pieces of evidence, the clearest one is the positive correlation between GABA levels and semantic task accuracy. However, it is important to note that this correlation already exists before the stimulation, and there are no results supporting that it can be modulated by the stimulation. In fact, cTBS significantly increases GABA levels but does not significantly improve performance on semantic tasks. According to the authors' interpretation of the results in Table 1, cTBS stimulation may have masked the practice effects that were supposed to occur. In other words, the stimulation decreased rather than enhanced participants' behavioral performance on the semantic task.The stimulation effect on behavioral performance could potentially be explained by the nonlinear relationship between GABA and performance on semantic tasks proposed by the authors. However, the current results are also insufficient to support the authors' hypothesis of an inverted U-shaped curve. Firstly, in Figure 3C and Figure 3D, the last one-third of the inverted U-shaped curve does not have any data points. In other words, as the GABA level increases the accuracy of the behavior first rises and then remains at a high level. This pattern of results may be due to the ceiling effect of the behavioral task's accuracy, rather than an inverted U-shaped ATL GABA function in semantic memory. Second, the article does not provide sufficient evidence to support the existence of an optimal level of GABA in the ATL. Fortunately, this can be tested with additional data analysis. The authors can estimate, based on pre-stimulus data from individuals, the optimal level of GABA for semantic functioning. They can then examine two expectations: first, participants with pre-stimulus GABA levels below the optimal level should show improved behavioral performance after stimulation-induced GABA elevation; second, participants with pre-stimulus GABA levels above the optimal level should exhibit a decline in behavioral performance after stimulation-induced GABA elevation. Alternatively, the authors can categorize participants into groups based on whether their behavioral performance improves or declines after stimulation, and compare the pre- and post-stimulus GABA levels between the two groups. If the improvement group shows significantly lower pre-stimulus GABA levels compared to the decline group, and both groups exhibit an increase in GABA levels after stimulation, this would also provide some support for the authors' hypothesis.
Another issue in this study is the confounding of simulation effects and practice effects. According to the results, there is a significant improvement in performance after the simulation, at least in the control task, which the authors suggest may reflect a practice effect. The authors argue that the results in Table 1 suggest a similar practice effect in the semantic task, but it is masked by the simulation of the ATL. However, since no significant effects were found in the ANOVA analysis of the semantic task, it is actually difficult to draw a conclusion. This potential confound increases the risk in data analysis and interpretation. Specifically, for Figure 3D, if practice effects are taken into account, the data before and after the simulation should not be analyzed together.
-
Reviewer #2 (Public Review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data from their previous MRS study suggest an inverted …Reviewer #2 (Public Review):
Summary:
The authors combined inhibitory neurostimulation (continuous theta-burst stimulation, cTBS) with subsequent MRI measurements to investigate the impact of inhibition of the left anterior temporal lobe (ATL) on task-related activity and performance during a semantic task and link stimulation-induced changes to the neurochemical level by including MR spectroscopy (MRS). cTBS effects in the ATL were compared with a control site in the vertex. The authors found that relative to stimulation of the vertex, cTBS significantly increased the local GABA concentration in the ATL. cTBS also decreased task-related semantic activity in the ATL and potentially delayed semantic task performance by hindering a practice effect from pre to post. Finally, pooled data from their previous MRS study suggest an inverted U-shape between GABA concentration and behavioral performance. These results help to better understand the neuromodulatory effects of non-invasive brain stimulation on task performance.Strengths:
Multimodal assessment of neurostimulation effects on the behavioral, neurochemical, and neural levels. In particular, the link between GABA modulation and behavior is timely and potentially interesting.Weaknesses:
The analyses are not sound. Some of the effects are very weak and not all conclusions are supported by the data since some of the comparisons are not justified. There is some redundancy with a previous paper by the same authors, so the novelty and contribution to the field are overall limited. A network approach might help here. -
Reviewer #3 (Public Review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on …Reviewer #3 (Public Review):
Summary:
The authors used cTBS TMS, magnetic resonance spectroscopy (MRS), and functional magnetic resonance imaging (fMRI) as the main methods of investigation. Their data show that cTBS modulates GABA concentration and task-dependent BOLD in the ATL, whereby greater GABA increase following ATL cTBS showed greater reductions in BOLD changes in ATL. This effect was also reflected in the performance of the behavioural task response times, which did not subsume to practice effects after AL cTBS as opposed to the associated control site and control task. This is in line with their first hypothesis. The data further indicates that regional GABA concentrations in the ATL play a crucial role in semantic memory because individuals with higher (but not excessive) GABA concentrations in the ATLs performed better on the semantic task. This is in line with their second prediction. Finally, the authors conducted additional analyses to explore the mechanistic link between ATL inhibitory GABAergic action and semantic task performance. They show that this link is best captured by an inverted U-shaped function as a result of a quadratic linear regression model. Fitting this model to their data indicates that increasing GABA levels led to better task performance as long as they were not excessively low or excessively high. This was first tested as a relationship between GABA levels in the ATL and semantic task performance; then the same analyses were performed on the pre and post-cTBS TMS stimulation data, showing the same pattern. These results are in line with the conclusions of the authors.Strengths:
I thoroughly enjoyed reading the manuscript and appreciate its contribution to the field of the role of the ATL in semantic processing, especially given the efforts to overcome the immense challenges of investigating ATL function by neuroscientific methods such as MRS, fMRI & TMS. The main strengths are summarised as follows:• The work is methodologically rigorous and dwells on complex and complementary multimethod approaches implemented to inform about ATL function in semantic memory as reflected in changes in regional GABA concentrations. Although the authors previously demonstrated a negative relationship between increased GABA levels and BOLD signal changes during semantic processing, the unique contribution of this work lies within evidence on the effects of cTBS TMS over the ATL given by direct observations of GABA concentration changes and further exploring inter-individual variability in ATL neuroplasticity and consequent semantic task performance.
• Another major asset of the present study is implementing a quadratic regression model to provide insights into the non-linear relationship between inhibitory GABAergic activity within the ATLs and semantic cognition, which improves with increasing GABA levels but only as long as GABA levels are not extremely high or low. Based on this finding, the authors further pinpoint the role of inter-individual differences in GABA levels and cTBS TMS responsiveness, which is a novel explanation not previously considered (according to my best knowledge) in research investigating the effect of TMS on ATLs.
• There are also many examples of good research practice throughout the manuscript, such as the explicitly stated exploratory analyses, calculation of TMS electric fields, using ATL optimised dual echo fRMI, links to open source resources, and a part of data replicates a previous study by Jung et. al (2017).
Weaknesses:
• Research on the role of neurotransmitters in semantic memory is still very rare and therefore the manuscript would benefit from more context on how GABA contributes to individual differences in cognition/behaviour and more justification on why the focus is on semantic memory. A recommendation to the authors is to highlight and explain in more depth the particular gaps in evidence in this regard.• The focus across the experiments is on the left ATL; how do the authors justify this decision? Highlighting the justification for this methodological decision will be important, especially given that a substantial body of evidence suggests that the ATL should be involved in semantics bilaterally (e.g. Hoffman & Lambon Ralph, 2018; Lambon Ralph et al., 2009; Rice et al., 2017; Rice, Hoffman, et al., 2015; Rice, Ralph, et al., 2015; Visser et al., 2010).
• When describing the results, (Pg. 11; lines 233-243), the authors first show that the higher the BOLD signal intensity in ATL as a response to the semantic task, the lower the GABA concentration. Then, they state that individuals with higher GABA concentrations in the ATL perform the semantic task better. Although it becomes clearer with the exploratory analysis described later, at this point, the results seem rather contradictory and make the reader question the following: if increased GABA leads to less task-induced ATL activation, why at this point increased GABA also leads to facilitating and not inhibiting semantic task performance? It would be beneficial to acknowledge this contradiction and explain how the following analyses will address this discrepancy.
• There is an inconsistency in reporting behavioural outcomes from the performance on the semantic task. While experiment 1 (cTBS modulates regional GANA concentrations and task-related BOLD signal changes in the ATL) reports the effects of cTBS TMS on response times, experiment 2 (Regional GABA concentrations in the ATL play a crucial role in semantic memory) and experiment 3 (The inverted U-shaped function of ATL GABA concentration in semantic processing) report results on accuracy. For full transparency, the manuscript would benefit from reporting all results (either in the main text or supplementary materials) and providing further explanations on why only one or the other outcome is sensitive to the experimental manipulations across the three experiments.
Overall, the most notable impact of this work is the contribution to a better understanding of individual differences in semantic behaviour and the potential to guide therapeutic interventions to restore semantic abilities in neurological populations. While I appreciate that this is certainly the case, I would be curious to read more about how this could be achieved.
-