Deficiency of orexin receptor type 1 in dopaminergic neurons increases novelty-induced locomotion and exploration
Curation statements for this article:-
Curated by eLife

eLife Assessment
This manuscript describes valuable findings regarding the expression pattern of orexin receptors in the midbrain and how manipulating this system influences several behaviors, such as context-induced locomotor activity and exploration. The overall strength of evidence - which includes anatomical, viral manipulation studies, and brain imaging - is solid and broadly substantiates claims in the paper. However, there are several areas in which the conclusions are only partially supported by the combination of methods used. These results have implications for understanding the neural underpinnings of reward and will be of interest to neuroscientists and cognitive scientists with an interest in the neurobiology of reward.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Orexin signaling in the ventral tegmental area and substantia nigra promotes locomotion and reward processing, but it is not clear whether dopaminergic neurons directly mediate these effects. We show that dopaminergic neurons in these areas mainly express orexin receptor subtype 1 (Ox1R). In contrast, only a minor population in the medial ventral tegmental area express orexin receptor subtype 2 (Ox2R). To analyze the functional role of Ox1R signaling in dopaminergic neurons, we deleted Ox1R specifically in dopamine transporter-expressing neurons of mice and investigated the functional consequences. Deletion of Ox1R increased locomotor activity and exploration during exposure to novel environments or when intracerebroventricularely injected with orexin A. Spontaneous activity in home cages, anxiety, reward processing, and energy metabolism did not change. Positron emission tomography imaging revealed that Ox1R signaling in dopaminergic neurons affected distinct neural circuits depending on the stimulation mode. In line with an increase of neural activity in the lateral paragigantocellular nucleus (LPGi) of Ox1R ΔDAT mice, we found that dopaminergic projections innervate the LPGi in regions where the inhibitory dopamine receptor subtype D2 but not the excitatory D1 subtype resides. These data suggest a crucial regulatory role of Ox1R signaling in dopaminergic neurons in novelty-induced locomotion and exploration.
Article activity feed
-

-
-
-

eLife Assessment
This manuscript describes valuable findings regarding the expression pattern of orexin receptors in the midbrain and how manipulating this system influences several behaviors, such as context-induced locomotor activity and exploration. The overall strength of evidence - which includes anatomical, viral manipulation studies, and brain imaging - is solid and broadly substantiates claims in the paper. However, there are several areas in which the conclusions are only partially supported by the combination of methods used. These results have implications for understanding the neural underpinnings of reward and will be of interest to neuroscientists and cognitive scientists with an interest in the neurobiology of reward.
-
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay between these two systems in regulating behaviour by first characterizing the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, Dat-Cre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons but does not seem to induce c-Fos expression. Behavioral effects of depleting OX1R from dopaminergic neurons include enhanced novelty-induced locomotion and exploration, relative to littermate controls (Ox1R fl/fl, Dat-Cre wt/wt). However, no difference between groups is observed …
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay between these two systems in regulating behaviour by first characterizing the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, Dat-Cre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons but does not seem to induce c-Fos expression. Behavioral effects of depleting OX1R from dopaminergic neurons include enhanced novelty-induced locomotion and exploration, relative to littermate controls (Ox1R fl/fl, Dat-Cre wt/wt). However, no difference between groups is observed in tests that measure reward processing, anxiety, and energy homeostasis. To test whether the depletion of OX1R alters overall orexin-triggered activation across the brain, PET imaging is used in OX1R∆DAT knockout and control mice. This analysis reveals that several regions show higher neuronal activation after orexin injection in OX1R∆DAT mice, but the authors focus their follow-up study on the dorsal bed nucleus of the stria terminalis (BNST) and lateral paragigantocellular nucleus (LPGi). Dopaminergic inputs and expression of dopamine receptors type-1 and -2 (DRD1 & DRD2) are assessed and compared to control demonstrating a moderate decrease in DRD1 and DRD2 expression in the BNST of OX1R∆DAT mice and unaltered expression of DRD2, with absence of DRD1 expression in LPGi of both groups. Overall, this study is valuable for the information it provides on orexin receptor expression and function in behaviour, as well as for the new tools it generated for the specific study of this receptor in dopaminergic circuits.
Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter expressing neurons is a strong approach to dissect the direct role of orexin in modulating dopamine signaling in the brain. The battery of behavioral assays used to study this line provides valuable information for researchers interested in the interplay between dopamine and orexin systems and their role in animal physiology.
Weaknesses:
This study falls short in providing evidence for an anatomical substrate and mechanism underlying the altered behavior observed in mice lacking orexin receptor subtype 1 in dopaminergic neurons. How orexin transmission in dopaminergic neurons regulates the expression of postsynaptic dopamine receptors (as observed in the BNST of OX1R∆DAT mice) is an intriguing question not addressed in this study. An important aspect not investigated in this study is whether the disruption of orexin activity affects dopamine release in target areas.
-
Reviewer #2 (Public review):
Summary:
This manuscript examines expression of orexin receptors in midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that substantia nigra dopamine neurons predominantly express orexin receptor 1 subtype and then go on to delete this receptor in dopamine transporter-expressing neurons using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that, in the absence of this receptor, orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, …
Reviewer #2 (Public review):
Summary:
This manuscript examines expression of orexin receptors in midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that substantia nigra dopamine neurons predominantly express orexin receptor 1 subtype and then go on to delete this receptor in dopamine transporter-expressing neurons using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that, in the absence of this receptor, orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and exploration, anxiety-related behavior, preference for sweet solutions, cocaine-induced conditioned place preference, and energy metabolism. Of these, the most consistent effects are seen in the tests of novelty-induced locomotion and exploration in which the mice with orexin 1 receptor deletion are observed to show greater levels of exploration, relative to wild-type, when placed in a novel environment, an effect that is augmented after icv administration of orexin.
In the final part of the paper, the authors use PET imaging to compare brain-wide activity patterns in the mutant mice compared to wildtype. They find differences in several areas both under control conditions (i.e., after injection of saline) as well as after injection of orexin. They focus in on changes in dorsal bed nucleus of stria terminalis (dBNST) and the lateral paragigantocellular nucleus (LPGi) and perform analysis of the dopaminergic projections to these areas. They provide anatomical evidence that these regions are innervated by dopamine fibers from midbrain, are activated by orexin in control, but not mutant mice, and that dopamine receptors are present. They also show changes in receptor expression in the transgenic mice. Thus, they argue these anatomical data support the hypothesis that behavioral effects of orexin receptor 1 deletion in dopamine neurons are due to changes in dopamine signaling in these areas.
Strengths:
Understanding how orexin interacts with the dopamine system is an important question and this paper contains several novel findings along these lines. Specifically:
(1) Distribution of orexin receptor subtypes in VTA and SN is explored thoroughly.
(2) Use of the genetic model that knocks out a specific orexin receptor subtype from dopamine-transporter-expressing neurons is a useful model and helps to narrow down the behavioral significance of this interaction.
(3) PET studies showing how central administration of orexin evokes dopamine release across the brain is intriguing, especially since two key areas are pursued - BNST and LPGi - where the dopamine projection is not as well described/understood.Weaknesses:
The role of the orexin-dopamine interaction is not explored in enough detail. The manuscript presents several related findings, but the combination of anatomy and manipulation studies do not quite tell a cogent story. Ideally, one would like to see the authors focus on a specific behavioral parameter and show that one of their final target areas (dBNST or LPGi) was responsible or at least correlated with this behavioral readout. In addition, the authors' working model for how they think orexin-dopamine interactions contribute to behavior under normal physiological conditions is not well-described.
-
Author response:
The following is the authors’ response to the previous reviews.
Public Reviews:
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay of these two systems in regulating behaviour by first characterising the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, DatCre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons, but does not seem to induce c-Fos expression. Behavioural effects of depleting OX1R from dopaminergic neurons includes enhanced noveltyinduced locomotion and exploration, relative to …
Author response:
The following is the authors’ response to the previous reviews.
Public Reviews:
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay of these two systems in regulating behaviour by first characterising the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, DatCre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons, but does not seem to induce c-Fos expression. Behavioural effects of depleting OX1R from dopaminergic neurons includes enhanced noveltyinduced locomotion and exploration, relative to littermate controls (Ox1R fl/fl, Dat-Cre wt/wt). However, no difference between groups is observed in tests that measure reward processing, anxiety, and energy homeostasis. To test whether depletion of OX1R alters overall orexin-triggered activation across the brain, PET imaging is used in OX1R∆DAT knockout and control mice. This analysis reveals that several regions show a higher neuronal activation after orexin injection in OX1R∆DAT mice, but the authors focus their follow up study on the dorsal bed nucleus of the stria terminalis (BNST) and lateral paragigantocellular nucleus (LPGi). Dopaminergic inputs and expression of dopamine receptors type-1 and -2 (DRD1 & DRD2) is assessed and compared to control demonstrating moderate decrease of DRD1 and DRD2 expression in BNST of OX1R∆DAT mice and unaltered expression of DRD2, with absence of DRD1 expression in LPGi of both groups. Overall, this study is valuable for the information it provides on orexin receptor expression and function on behaviour and for the new tools it generated for the specific study of this receptor in dopaminergic circuits.
Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter expressing neurons is a strong approach to dissect the direct role of orexin in modulating dopamine signalling in the brain. The battery of behavioural assays to study this line provides a valuable source of information for researchers interested in the role of orexin in animal physiology.
We thank the reviewer for summarizing the importance and significance of our study.
Weaknesses:
This study falls short in providing evidence for an anatomical substrate of the altered behaviour observed in mice lacking orexin receptor subtype 1 in dopaminergic neurons. How orexin transmission in dopaminergic neurons regulates the expression of postsynaptic dopamine receptors (as observed in BNST of OX1R∆DAT mice) is an intriguing question poorly discussed. Whether disruption of orexin activity alters dopamine release in target areas is an important point not addressed.
We identified dopaminergic fibers and dopamine receptors in the dBNST and LPGi, suggesting anatomical basis for dopamine neurons to regulate neural activity and receptor expression levels in these areas. PET imaging scan and c-Fos staining revealed that Ox1R signaling in dopaminergic cells regulates neuronal activity in dBNST and LPGi. The expression levels of Th were unchanged in both regions. Dopamine receptor 2 (DRD2), but not DRD1, is expressed in LPGi. The deletion of Ox1R in DAT-expressing cells did not affect DRD2 expression in LPGi. The expression levels of DRD1 and DRD2 were decreased or showed a tendency to decrease in dBNST.
We included the comments in the discussion in this revised manuscript (lines 308-312): ‘The expression levels of Th were not altered in dBNST or LPGi by Ox1R deletion in dopaminergic neurons. It remains unclear whether dopamine release is affected in these regions. It is possible that either the dopaminergic regulation of neuronal activity or the changes in dopamine release could lead to the decreased expression of dopamine receptors in dBNST.’
Reviewer #2 (Public review):
Summary:
This manuscript examines expression of orexin receptors in midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express orexin receptor 1 subtype and then go on to delete this receptor in dopamine transporter-expressing using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that, in the absence of this receptor, orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and exploration, anxiety-related behavior, preference for sweet solutions, cocaine-induced conditioned place preference, and energy metabolism. Of these, the most consistent effects are seen in the tests of novelty-induced locomotion and exploration in which the mice with orexin 1 receptor deletion are observed to show greater levels of exploration, relative to wild-type, when placed in a novel environment, an effect that is augmented after icv administration of orexin.
In the final part of the paper, the authors use PET imaging to compare brain-wide activity patterns in the mutant mice compared to wildtype. They find differences in several areas both under control conditions (i.e., after injection of saline) as well as after injection of orexin. They focus in on changes in dorsal bed nucleus of stria terminalis (dBNST) and the lateral paragigantocellular nucleus (LPGi) and perform analysis of the dopaminergic projections to these areas. They provide anatomical evidence that these regions are innervated by dopamine fibers from midbrain, are activated by orexin in control, but not mutant mice, and that dopamine receptors are present. Thus, they argue these anatomical data support the hypothesis that behavioral effects of orexin receptor 1 deletion in dopamine neurons are due to changes in dopamine signaling in these areas.
Strengths:
Understanding how orexin interacts with the dopamine system is an important question and this paper contains several novel findings along these lines. Specifically:
(1) Distribution of orexin receptor subtypes in VTA and SN is explored thoroughly.
(2) Use of the genetic model that knocks out a specific orexin receptor subtype from dopaminetransporter-expressing neurons is a useful model and helps to narrow down the behavioral significance of this interaction.
(3) PET studies showing how central administration of orexin evokes dopamine release across the brain is intriguing, especially that two key areas are pursued - BNST and LPGi - where the dopamine projection is not as well described/understood.
We thank the reviewer for summarizing the importance and significance of our study.
Weaknesses:
The role of the orexin-dopamine interaction is not explored in enough detail. The manuscript presents several related findings, but the combination of anatomy and manipulation studies do not quite tell a cogent story. Ideally, one would like to see the authors focus on a specific behavioral parameter and show that one of their final target areas (dBNST or LPGi) was responsible or at least correlated with this behavioral readout.
We agree that exploring the orexin-dopamine interactions in more detail and focusing on the behavioral impact of their final target areas (e.g., dBNST or LPGi), would provide valuable data. While we are very interested in pursuing these studies, the aim of the present manuscript is to provide an overview of the behavioral roles of orexin-dopamine interaction and to propose some promising downstream pathways in a relatively broad and systematic manner.
In many places in the Results, insufficient explanation and statistical reporting is provided. Throughout the Results - especially in the section on behavior although not restricted to this part - statements are made without statistical tests presented to back up the claims, e.g., "Compared to controls, Ox1RΔDAT 143 mice did not show significant changes in spontaneous locomotor activity in home cages" (L143) and "In a hole-board test, female Ox1RΔDAT mice showed increased nose pokes into the holes in early (1st and 2nd) sessions compared to control mice" (L151). In other places, ANOVAs are mentioned but full results including main effects and interactions are not described in detail, e.g., in F3-S3, only a single p-value is presented and it is difficult to know if this is the interaction term or a post hoc test (L205). These and all other statements need statistics included in the text as support. Addition of these statistical details was also requested by the editor.
We submitted all our source data as Excel spreadsheets to eLife during our first-round revision, and the full statistics, such as main effects and interactions, are presented alongside the source data in the respective spreadsheets. We thank the reviewer for pointing out our lack of clarity in the manuscript. In this revised manuscript, we included the statistical details of ANOVAs mentioned above in the figure legends. In the figure legends, we also explained that the full statistics were provided alongside the source data in the supplementary materials.
In the presentation of reward processing this is particularly important as no statistical tests are shown to demonstrate that controls show a cocaine-induced preference or a sucrose preference. Here, one option would be to perform one-sample t-tests showing that the data were different to zero (no preference). As it is, the claim that "Both of the control and Ox1RΔDAT groups showed a preference for cocaine injection" is not yet statistically supported.
We thank the reviewer for the suggestions. We have added the one-sample t-test results in this revised manuscript (Figure 2–figure supplement 4, lines 171 - 183).
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
Can the authors comment on overlap between DAT and Ox1R in brain areas outside VTA/SN? Is there any?
We only focused on the expression patterns of orexin receptors in VTA/SN, and we did not examine other brain regions. Additionally, little is known from the literature about the expression of Ox1R in DAT-expressing cells in brain areas outside VTA/SN. Further analysis is necessary to answer this question. We have added the comment in our discussion (lines 243 - 344).
For the Ca2+ imaging experiment, it is unclear to me why the authors do not show all the neurons (almost 160 in total) and just select 5 neurons to show for each condition.
Heat maps of all recorded neurons are now shown in Figure 1—figure supplement 4.
There are other claims that still require a statistical justification to be included in addition to the passages on behavior mentioned above, e.g., "Increasing the orexin A concentration to 300 nM further increased [Ca2+]i" (L118).
Authors should ensure that all such claims are either presented with a statistical test or are phrased differently, e.g. "Visual inspection of data suggested that there was a further increase...". In addition, when an ANOVA is conducted, full results including main effects and interactions should be described.
We emphasize now our statement that ALREADY 100 nM orexin A significantly increased [Ca2+]i levels (lines 117 - 118).
We submitted all our source data as Excel spreadsheets to eLife during our first-round revision, and the full statistics, such as main effects and interactions, are presented alongside the source data in the respective spreadsheets. For clarity, we chose to include only the key statistical information in the main text and figures. We thank the reviewer for pointing this out. In this revised manuscript, we have emphasized in each figure legend: ‘Source data and full statistics are provided in the supplementary materials’.
Typos in figure captions
F2-S1 - spontanous
F3-S2 - intrest
We apologize for the typos. We have corrected them in this revised manuscript.
Editor's note:
Should you choose to revise your manuscript, please include full statistical reporting including exact p-values wherever possible alongside the summary statistics (test statistic and df) and 95% confidence intervals. These should be reported for all key questions and not only when the p-value is less than 0.05.
We submitted all our source data as Excel spreadsheets to eLife during our first-round revision, and the full statistics, such as test statistics, df and 95% confidence intervals, are presented alongside the source data in the respective spreadsheets. We thank the editor’s note. In this revised manuscript, we have included more statistical information in the main text and figure legends (see our response to reviewer #2). In the figure legends, we also explained that the full statistics were provided alongside the source data in the supplementary materials. In addition, we also uploaded the source data and full statistics in the bioRxiv before we upload this revised manuscript to eLife.
-
-
eLife assessment
This manuscript describes valuable findings on the expression pattern of orexin receptors in the midbrain and how manipulating this system influences several behaviors, such as context-induced locomotor activity and exploration. The overall strength of evidence - which includes anatomical, viral manipulation studies, and brain imaging - is solid and broadly supports claims in the paper, however, there are several areas in which the conclusions are only partially supported by the statistical evidence. These results have implications for understanding the neural underpinnings of reward and will be of interest to neuroscientists and cognitive scientists with an interest in the neurobiology of reward.
-
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay of these two systems in regulating behaviour by first characterising the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, Dat-Cre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons, but does not seem to induce c-Fos expression. Behavioural effects of depleting OX1R from dopaminergic neurons includes enhanced novelty-induced locomotion and exploration, relative to littermate controls (Ox1R fl/fl, Dat-Cre wt/wt). However, no difference between groups is observed in …
Reviewer #1 (Public review):
In this manuscript, the role of orexin receptors in dopamine transmission is studied. It extends previous findings suggesting an interplay of these two systems in regulating behaviour by first characterising the expression of orexin receptors in the midbrain and then disrupting orexin transmission in dopaminergic neurons by deleting its predominant receptor, OX1R (Ox1R fl/fl, Dat-Cre tg/wt mice). Electrophysiological and calcium imaging data suggest that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons, but does not seem to induce c-Fos expression. Behavioural effects of depleting OX1R from dopaminergic neurons includes enhanced novelty-induced locomotion and exploration, relative to littermate controls (Ox1R fl/fl, Dat-Cre wt/wt). However, no difference between groups is observed in tests that measure reward processing, anxiety, and energy homeostasis. To test whether depletion of OX1R alters overall orexin-triggered activation across the brain, PET imaging is used in OX1R∆DAT knockout and control mice. This analysis reveals that several regions show a higher neuronal activation after orexin injection in OX1R∆DAT mice, but the authors focus their follow up study on the dorsal bed nucleus of the stria terminalis (BNST) and lateral paragigantocellular nucleus (LPGi). Dopaminergic inputs and expression of dopamine receptors type-1 and -2 (DRD1 & DRD2) is assessed and compared to control demonstrating moderate decrease of DRD1 and DRD2 expression in BNST of OX1R∆DAT mice and unaltered expression of DRD2, with absence of DRD1 expression in LPGi of both groups. Overall, this study is valuable for the information it provides on orexin receptor expression and function on behaviour and for the new tools it generated for the specific study of this receptor in dopaminergic circuits.
Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter expressing neurons is a strong approach to dissect the direct role of orexin in modulating dopamine signalling in the brain. The battery of behavioural assays to study this line provides a valuable source of information for researchers interested in the role of orexin in animal physiology.
Weaknesses:
This study falls short in providing evidence for an anatomical substrate of the altered behaviour observed in mice lacking orexin receptor subtype 1 in dopaminergic neurons. How orexin transmission in dopaminergic neurons regulates the expression of postsynaptic dopamine receptors (as observed in BNST of OX1R∆DAT mice) is an intriguing question poorly discussed. Whether disruption of orexin activity alters dopamine release in target areas is an important point not addressed.
-
Reviewer #2 (Public review):
Summary:
This manuscript examines expression of orexin receptors in midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express orexin receptor 1 subtype and then go on to delete this receptor in dopamine transporter-expressing using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that, in the absence of this receptor, orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion …
Reviewer #2 (Public review):
Summary:
This manuscript examines expression of orexin receptors in midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express orexin receptor 1 subtype and then go on to delete this receptor in dopamine transporter-expressing using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that, in the absence of this receptor, orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and exploration, anxiety-related behavior, preference for sweet solutions, cocaine-induced conditioned place preference, and energy metabolism. Of these, the most consistent effects are seen in the tests of novelty-induced locomotion and exploration in which the mice with orexin 1 receptor deletion are observed to show greater levels of exploration, relative to wild-type, when placed in a novel environment, an effect that is augmented after icv administration of orexin.
In the final part of the paper, the authors use PET imaging to compare brain-wide activity patterns in the mutant mice compared to wildtype. They find differences in several areas both under control conditions (i.e., after injection of saline) as well as after injection of orexin. They focus in on changes in dorsal bed nucleus of stria terminalis (dBNST) and the lateral paragigantocellular nucleus (LPGi) and perform analysis of the dopaminergic projections to these areas. They provide anatomical evidence that these regions are innervated by dopamine fibers from midbrain, are activated by orexin in control, but not mutant mice, and that dopamine receptors are present. Thus, they argue these anatomical data support the hypothesis that behavioral effects of orexin receptor 1 deletion in dopamine neurons are due to changes in dopamine signaling in these areas.
Strengths:
Understanding how orexin interacts with the dopamine system is an important question and this paper contains several novel findings along these lines. Specifically:
(1) Distribution of orexin receptor subtypes in VTA and SN is explored thoroughly.
(2) Use of the genetic model that knocks out a specific orexin receptor subtype from dopamine-transporter-expressing neurons is a useful model and helps to narrow down the behavioral significance of this interaction.
(3) PET studies showing how central administration of orexin evokes dopamine release across the brain is intriguing, especially that two key areas are pursued - BNST and LPGi - where the dopamine projection is not as well described/understood.Weaknesses:
The role of the orexin-dopamine interaction is not explored in enough detail. The manuscript presents several related findings, but the combination of anatomy and manipulation studies do not quite tell a cogent story. Ideally, one would like to see the authors focus on a specific behavioral parameter and show that one of their final target areas (dBNST or LPGi) was responsible or at least correlated with this behavioral readout.
In many places in the Results, insufficient explanation and statistical reporting is provided. Throughout the Results - especially in the section on behavior although not restricted to this part - statements are made without statistical tests presented to back up the claims, e.g., "Compared to controls, Ox1RΔDAT 143 mice did not show significant changes in spontaneous locomotor activity in home cages" (L143) and "In a hole-board test, female Ox1RΔDAT mice showed increased nose pokes into the holes in early (1st and 2nd) sessions compared to control mice" (L151). In other places, ANOVAs are mentioned but full results including main effects and interactions are not described in detail, e.g., in F3-S3, only a single p-value is presented and it is difficult to know if this is the interaction term or a post hoc test (L205). These and all other statements need statistics included in the text as support. Addition of these statistical details was also requested by the editor.
In the presentation of reward processing this is particularly important as no statistical tests are shown to demonstrate that controls show a cocaine-induced preference or a sucrose preference. Here, one option would be to perform one-sample t-tests showing that the data were different to zero (no preference). As it is, the claim that "Both of the control and Ox1RΔDAT groups showed a preference for cocaine injection" is not yet statistically supported.
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
In this manuscript, the role of orexin receptors in dopamine neurons is studied. Considering the importance of both orexin and dopamine signalling in the brain, with critical roles in arousal and drug seeking, this study is important to understand the anatomical and functional interaction between these two neuromodulators. This work suggests that such interaction is direct and occurs at the level of SN and VTA, via the expression of OX1R-type orexin receptors by dopaminergic neurons.
Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter-expressing neurons is a strong approach to dissecting the direct role of orexin in modulating dopamine signalling in the brain. The battery of …
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
In this manuscript, the role of orexin receptors in dopamine neurons is studied. Considering the importance of both orexin and dopamine signalling in the brain, with critical roles in arousal and drug seeking, this study is important to understand the anatomical and functional interaction between these two neuromodulators. This work suggests that such interaction is direct and occurs at the level of SN and VTA, via the expression of OX1R-type orexin receptors by dopaminergic neurons.
Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter-expressing neurons is a strong approach to dissecting the direct role of orexin in modulating dopamine signalling in the brain. The battery of behavioural assays to study this line provides a valuable source of information for researchers interested in the role of orexin-A in animal physiology.
We thank the reviewer for summarizing the importance and significance of our study.
Weaknesses:
The choice of methods to demonstrate the role of orexin in the activation of dopamine neurons is not justified and the quantification methods are not described with enough detail. The representation of results can be dramatically improved and the data can be statistically analysed with more appropriate methods.
We have further improved our description of the methods in the revised reviewed preprint, and here in the response letter, we respond point-by-point to ‘Reviewer #1 (Recommendations For The Authors)’ below.
Reviewer #2 (Public Review):
Summary:
This manuscript examines the expression of orexin receptors in the midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express the orexin receptor 1 subtype and then go on to delete this receptor in dopamine neurons using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that in the absence of this receptor orexin is no longer able to excite dopamine neurons of the substantia nigra.
The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and exploration, anxiety-related behavior, preference for sweet solutions, cocaine-induced conditioned place preference, and energy metabolism. Of these, the most consistent effects are seen in the tests of novelty-induced locomotion and exploration in which the mice with orexin 1 receptor deletion are observed to show greater levels of exploration, relative to wild-type, when placed in a novel environment, an effect that is augmented after icv administration of orexin.
In the final part of the paper, the authors use PET imaging to compare brain-wide activity patterns in the mutant mice compared to wildtype. They find differences in several areas both under control conditions (i.e., after injection of saline) as well as after injection of orexin. They focus on changes in the dorsal bed nucleus of stria terminalis (dBNST) and the lateral paragigantocellular nucleus (LPGi) and perform analysis of the dopaminergic projections to these areas. They provide anatomical evidence that these regions are innervated by dopamine fibers from the midbrain, are activated by orexin in control, but not mutant mice, and that dopamine receptors are present. Thus, they argue these anatomical data support the hypothesis that behavioral effects of orexin receptor 1 deletion in dopamine neurons are due to changes in dopamine signaling in these areas.
Strengths:
Understanding how orexin interacts with the dopamine system is an important question and this paper contains several novel findings along these lines. Specifically:
(1) The distribution of orexin receptor subtypes in VTA and SN is explored thoroughly.
(2) Use of the genetic model that knocks out a specific orexin receptor subtype from only dopamine neurons is a useful model and helps to narrow down the behavioral significance of this interaction.
(3) PET studies showing how central administration of orexin evokes dopamine release across the brain is intriguing, especially since two key areas are pursued - BNST and LPGi - where the dopamine projection is not as well described/understood.
We thank the reviewer for the careful summary and highlighting the novelty of our study.
Weaknesses:
The role of the orexin-dopamine interaction is not explored in enough detail. The manuscript presents several related findings, but the combination of anatomy and manipulation studies does not quite tell a cogent story. Ideally, one would like to see the authors focus on a specific behavioral parameter and show that one of their final target areas (dBNST or LPGi) was responsible or at least correlated with this behavioral readout. In addition, some more discussion on what the results tell us about orexin signaling to dopamine neurons under normal physiological conditions would be very useful. For example, what is the relevance of the orexin-dopamine interaction blunting noveltyinduced locomotion under wildtype conditions?
We agree that focusing on some orexin-dopamine targeting areas, such as dBNST or LPGi, is important to further reveal the anatomy-behavior links and underlying mechanisms. While we are very interested in further investigations, in the present manuscript we mainly aim to give an overview of the behavioral roles of orexin-dopamine interaction and to propose some promising downstream pathways in a relatively broad and systematical way.
We have explained the physiological meanings of our results in more detail in the discussion in the revised reviewed preprint (lines 282-293, 318-332, ). Novelty-induced behavioral response should be at proper levels under normal physiological conditions. The orexin-dopamine interaction blunting novelty-induced locomotion could be important to keep attention on the main task without being distracted too much by other random stimuli in the environment. When this balance is disrupted, behavioral deficit may happen, such as attention deficit and hyperactivity disorder (ADHD).
In some places in the Results, insufficient explanation and reporting is provided. For example, when reporting the behavioral effects of the Ox1 deletion in two bottle preference, it is stated that "[mutant] mice showed significant changes..." without stating the direction in which preference was affected.
For the reward-related behaviors described in this study, we did not find significant changes between [mutant] and control mice. We agree that it will be helpful for readers by describing the behavioral tests in more details. In the revised reviewed preprint, we have described in more detail in the results and Materials and Methods section how the control and [mutant] mice behave to the reward (lines 162-165, 171-181, 526-528).
The cocaine CPP results are difficult to interpret because it is unclear whether any of the control mice developed a CPP preference. Therefore, it is difficult to conclude that the knockout animals were unaffected by drug reward learning. Similarly, the sucrose/sucralose preference scores are also difficult to interpret because no test of preference vs. water is performed (although the data appear to show that there is a preference at least at higher concentrations, it has not been tested).
We described the CPP analysis in the Materials and Methods section (lines 523-528 ) as below: ‘The percentage of time spent in the reward-paired compartment was calculated: 100 x time spent in the compartment / (total time - time spent in the middle area). The CPP score was then analyzed using the calculated percentage of time: 100 x (time on the test day – time on pre-test days)/ time on pre-test days. The pre-test and test days were before and after the conditioning, respectively. Thus, the CPP score above zero indicates that the CPP preference has developed.’ In Figure 2—figure supplement 4 C and F, it was shown that most control and knockout mice had a CPP score above zero. The control and knockout groups both developed a preference and there was no significant difference between the groups.
For the sucrose/sucralose preference tests, in Figure 2—figure supplement 4 A and D, we present values as the percentages of sucrose/sucralose consumption in total daily drinking amount (sucrose/sucralose solution + water). Thus, percentages above 50% indicates mice prefer sucrose/sucralose to water. As shown in the figure, male mice only showed weak preference of 0.5% sucrose, compared to water, and under all other tested conditions, the mice showed strong preference of the sweet solution. There was no significant difference between control and knockout mice.
We have described this in more details in the Results and Materials and Methods section in the revised reviewed preprint.
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
(1) Figure 1, A-I. It is difficult to depict the anatomical subdivision of VTA in Figure 1, panels A and B. It is recommended to add a panel showing a schematic illustration of the SNc and subregions of VTA: PN, PIF, PBP, IF (providing more detail than in Figure 1, panel J). It is also recommended to show lower magnification images (as in Figure 1 - supplement 1), including both hemispheres, and to delineate the outline of the different subregions using curved lines, based on reference atlases (similar to Figure 1, panel I, please include distance from bregma). It would be helpful to indicate in Figure 1 that panel A is a control mouse and panel B is a Ox1RΔDAT mouse and include C-F letters to show corresponding insets. Anatomically, the paraintrafasicular nucleus (PIF) is positioned between the paranigral nucleus (PN) and the parabrachial pigmented nucleus (PBP). The authors have depicted the PIF ventral to the PN in Figure 1 panels A, B, and I. These panels and the quantification of Ox1R/2R positive cells within the different subdivisions need to be corrected accordingly. The image analysis method used to quantify RNAscope fluorescent images is not described in sufficient detail. Please expand this section.
According to the reviewer’s suggestions, we have refined Figure 1 in the revised reviewed preprint. We are now showing the schematic illustration of the SN and subregions of VTA in panel I, with blue squares to label the regions shown in panels A and B, and the distance from bregma is included. The outlines to delineate SN and the subregions of VTA are adjusted from straight to curved lines based on reference atlases. As suggested, we have also indicated panel A is a control and panel B is a Ox1RΔDAT mouse and included C-F letters to show corresponding insets. We apologize for the mistake about labeling PIF and PN positions in Figure A. We have corrected the labeling of their positions and double checked the quantification accordingly. This does not change our discussion or conclusion since both PIF and PN are the medial part of VTA, where both Ox1R and Ox2R are observed. The description of the image analysis in Matierials and Methods section has been improved (lines 378-385). We decided not to show lower magnification images than in Figure 1—supplement 1 to include both hemispheres, in the interests of clarity and reader-friendliness.
(2) Figure 1, J-L. The claim that orexin activates dopaminergic SN and VTA neurons is weakly supported by the data provided. Calcium imaging of SN dopaminergic neurons in control mice suggests a discrete effect of 100 nM orexin-A application compared to baseline. Application of 300 nM shows a slightly bigger effect, but none of these results are statistically analysed.
We are surprised by this comment and thank the reviewer for pointing out our apparent lack of clarity in the previous version (lines 96-106 and legend of Figure 1K, L). In more detail, we explain the data analysis in the new version (lines 119-133, 451-465) and the legend of Figure 1K, L and Figure 1-figure supplement 3).
The main goal of this part of the project was to functionally validate the Ox1R knockout in dopaminergic (DAT-expressing) neurons. This was a prerequisite for the behavioral and PET imaging experiments. We used GCaMP-mediated Ca2+ imaging in acute brain slices to reach this goal. This analysis was performed on the dopaminergic SN neurons, which we used as an "indicator population" because a large number of these neurons express Ox1R, but only a few express Ox2R.
The analysis consisted of two parts:
a) For each neuron, we tested whether it responded to orexin A. At the single cell level, a neuron was considered orexin A-responsive if the change in fluorescence induced by orexin A was three times larger than the standard deviation (3 σ criterion) of the baseline fluorescence, corresponding to a Zscore of 3. We found that 56% of the neurons tested responded to orexin A, while 44% of the neurons did not respond to orexin A (Figure 1L, top). These data agree with the number of Ox1R-expressing neurons (Figure 1J).
b) We also determined the orexin A-induced GCaMP fluorescence for each neuron, expressed as a percentage of GCaMP fluorescence induced upon application of high K+ saline. Accordingly, the "population response" of all analyzed neurons was expressed as the mean ± SEM of these responses. The significance of this mean response was tested for each group (control and Ox1R KO) using a onesample t-test. We found a marked and highly significant (p < 0.0001, n = 71) response of control neurons to 100 nM orexin A, while the Ox1R KO neurons did not respond (p = 0.5, n = 86). Note that, as described in a), 44% of the neurons contributing to the mean do not respond to orexin. Thus, the orexin responses of most responders are significantly higher than the mean. This is also evident in the example recordings in Figure 1K and Figure1—figure supplement 3. The orexin A-induced change in fluorescence was increased by increasing the orexin A concentration to 300 nM.
Note: As mentioned above, the orexin A response was expressed for each neuron individually as a percentage of its high K+saline-induced GCaMP fluorescence. This value is a solid reference point, reflecting the GCaMP fluorescence at maximal voltage-activated Ca2+ influx. Obviously, the Ca2+ concentration at this point is extremely high and not typically reached under physiological conditions. Therefore, as shown in Figure1—figure supplement 3 for completeness, the physiologically relevant responses may appear relatively minor at first glance when presented together in one figure (compare Figure1—figure supplement 3 A and B).
The authors should provide more evidence of the orexin-induced activation of dopaminergic neurons in the SN to support this claim and investigate whether a similar activation is observed in VTA neurons.
Following the reviewer's suggestion, we confirmed orexin A-induced activation of dopaminergic neurons in the mouse SN by using perforated patch clamp recordings (Figure1—figure supplement 2).
This finding is consistent with previous extracellular in vivo recordings in rats (Liu et al., 2018).
The activation of dopaminergic neurons in the mouse VTA by orexin A has been shown repeatedly in earlier studies (e.g., Baimel et al., 2017; Korotkova et al., 2003; Tung et al., 2016).
In addition, Figure 3-Figure Supplement 2 shows that injection of orexin does not induce c-Fos expression in SN and VTA dopaminergic neurons of control and Ox1RΔDAT mice, which further weakens the claim made by the authors.
Figure 3—Figure Supplement 2 in the original submission is now Figure 3—Figure Supplement 3 in the revised reviewed preprint. It shows low c-Fos expression in SN and VTA dopaminergic neurons, and orexin-induced c-Fos expression was observed in Th-negative cells in SN and VTA.
Technically relatively straightforward, Fos analysis is widely (and successfully) used in studies to reveal neuronal activation. However, this approach has limitations, e.g., regarding sensitivity and temporal resolution. Electrophysiological or optical imaging techniques can circumvent these shortcomings. The electrophysiological and Ca2+ imaging studies presented here, along with previous electrophysiological studies by others, clearly show that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons.
In vivo, the injection of orexin A induced a pronounced c-Fos activity in non-dopaminergic cells of the VTA and SN but not in dopaminergic neurons. This result shows that the detection of c-Fos has worked in principle. Whether the absent c-Fos staining in dopaminergic neurons is due to lack of sensitivity, whether other IEGs would have worked better here, or whether there are other, e.g., cell type-specific reasons for the absence of staining, cannot be determined from the current data.
(3) Figure 2, I-L. The fact that ICV injection of both saline and orexin causes a sustained increase of locomotion (around 20 minutes in males, and over 30 minutes in females) is problematic and could mask the effects of orexin, particularly in females. It is unclear what panels J and L are showing. To be appropriately analysed, the authors should plot the pre- and post-injection AUC data for all groups and analyse it as a two-way mixed ANOVA, with the within-subjects factor "pre/post injection activity" and between-subjects factor "group". The authors can only warrant a statistically meaningful hyperlocomotor effect in Ox1RΔDAT mice if a significant interaction is found.
Though mice were habituated to the injection, it still makes sense to see the injection-induced increase in locomotion to some extent. We described in the figure legend that the AUC was calculated for the period after orexin injection, which meant 5 – 90 min in Figure 2 I, K. We have clearly observed significant differences between genotypes and between saline and orexin application, which means the genotype and orexin impact is strong enough to pop up despite of the injection effect.
As the reviewer’s suggests, we have now plotted the pre- and post-injection AUC data for all groups and analyzed it as a two-way mixed ANOVA, with the within-subjects factor "pre/post injection activity" and between-subjects factor "group". To match the pre- and post-injection duration, we are now comparing AUC for around 60 min before and after the injection. A significant interaction is found here. Panels I-L are renewed, and the differences induced by Ox1R knockout and orexin confirmed the results shown in the initially submitted manuscript.
(4) Figure 3. The literature has robustly shown that one of the main projection areas of VTA and SN dopaminergic neurons is the striatum, in particular its ventral part. It is surprising to see that this region is not affected by the lack of OX1R or by the injection of orexin. How can the authors explain that identified regions with significantly different activity include neighbouring brain structures with heterogenous composition? See for example, in panel A, section bregma 0.62mm, a significant region is seen expanding across the cortex, corpus callosum, and striatum. While the data from PET studies is potentially interesting, it may not be adequate to provide enough resolution to allow examination of the anatomical distribution of orexin-mediated neuronal activation.
While the striatum is a major projection area of dopaminergic neurons in VTA and SN, the projection and function of Ox1R-positive dopaminergic neurons is not clear. We have improved the description of dopamine function diversity in the revised reviewed preprint (lines 46-58), and it was reported before that the projection-defined dopaminergic populations in the VTA exhibited different responses to orexin A (Baimel et al., 2017). Moreover, the striatum activity is modulated by the indirect effect via other brain regions affected by Ox1R-positive dopaminergic neurons. It is unknown how the striatum activity should change after Ox1R deletion in dopaminergic neurons. We could not rule out the possibility that the striatum is indeed modulated by the Ox1R-positive dopaminergic neurons, though there was only a trend of genotype difference (Ox1RΔDAT vs. ctrl) in the ventral striatum in the section bregma 1.42 mm in Figure 3A. The ICV injection of orexin is potentially acting on Ox1R and Ox2R in the whole brain, so projections from other brain regions to the striatum also affect striatum activity and could have masked the effect of Ox1R-positive dopaminergic neurons.
The spatial resolution of the PET data is in the order of ~1 mm^3. As we also explained in the Materials and Methods section, the size of a voxel in the original PET data is 0.4mm x 0.4mm x 0.8 mm. All calculations were performed on this grid. The higher-resolved images shown in Figure 3 are for presentation purposes only inspired by a request of the reviewer who asked us to show this in the Jais et al. 2016 manuscript. To make this clearer we now added the p-map images with the original voxel size to the supplement (Figure 3—figure supplement 1). For the interest in specific brain areas, more precise identification of anatomical sub-regions requires using methods with higher spatial resolution such as staining of brain slices for c-Fos-positive cells as we do in Figure 4.
PET is a powerful tool to identify global regions of activation/inhibition. In the manuscript, we have described in the results and discussion section that the activity in brain regions with related functions were changed. In panel A, Ox1RΔDAT showed activity increase in MPA, Pir and endopiriform claustrum, which are important for olfactory sensation; spinal trigeminal nucleus, sp5, and IRt, which regulates mastication and sensation of the oral cavity and the surface of the face; SubCV and Gi, which regulates sleeping and motion-related arousal and motivation. In panel B, changes in HDB, MCPO, Pir, DEn, S1, V2L and V1 are related to sensation, and changes in BNST, LPGi and M2 are important for emotion, exploration, and action selection.
(5) Figure 4. As in Figure 1, the authors should consider including a schematic illustration of the brain areas that are being analysed using a reference atlas. It is also recommended to provide more details describing the quantification of the images. Without such information, the data is not convincing, in particular, the claim that Ox1R depletion causes a decrease in DRD1 in BNST is unclear. Additional unbiased quantitative approaches could be used to strengthen this point.
We have added Figure 4—figure supplement 1 as a schematic illustration of the brain areas that were being analyzed using a reference atlas. More details describing the unbiased quantification of the images have been added to Materials and Methods. We have added Figure 4—figure supplement 3, to show DRD1, DRD2 and the merged signal separately.
(6) The discussion starts by stating that the main findings of this study are based on RNAscope and optophysiological experiments, however, the latter are not presented anywhere in the manuscript. This sentence (line 192) should be revised. The authors state in line 193 that OX1R is the only orexin receptor in the SN, but they show in Figure 1 that in the SN, 3% of neurons express OX2R and 2% co-express both receptors.
We thank the reviewer for the input. We have rephrased the beginning of the discussion to clarify the objectives (lines 238 - 246). In doing so, we changed "optophysiological experiments" and "single orexin receptor" (lines 192 and 193 in the original manuscript) to " Ca2+ imaging experiments" and "main subtype of orexin receptors ", respectively. In this context, it should be noted that Ca2+ imaging is considered an optophysiological method - optophysiology generally refers to techniques that combine optical methods with physiological measurements.
The results of LPGi and BNST dopamine receptors in control and Ox1RΔDAT mice are poorly discussed. The authors should justify why these two regions were selected for further validation and how these may be related to the behavioural effects found in Ox1RΔDAT regarding exposure to a novel context.
Ox1RΔDAT mice exhibited increased novelty- and orexin-induced locomotion compared to control mice. After orexin injection, PET imaging shows that the neural activity of BNST and LPGi was lower or higher than in control mice, respectively. We selected BNST and LPGi for further validation because we think their key functional roles in regulating emotion, exploratory behaviors and locomotor speed are related to novelty-induced locomotion. We confirmed changes in neural activity change by c-Fos staining and investigated the expression patterns of dopamine receptors in BNST and LPGi. Our findings suggested that Ox1R deletion in dopaminergic neurons results in the disinhibition of neural activity in LPGi via dopaminergic pathways and the decrease of dopamine-mediated neural activity in BNST. Emotion perception affects the decision of how to respond to the novelty. It is possible that novelty activates the orexin system and Ox1R signaling in dopaminergic neurons promotes emotion perception and inhibits exploration. Of course, further careful investigation is necessary to test this hypothesis in the future experiments. We have improved the rational description and discussion in the
‘Results’ and ‘Discussion’ section in the revised reviewed preprint (lines 210-213, 259-270, 293-308).
Reviewer #2 (Recommendations For The Authors):
A major recommendation - if possible - would be to directly show that one or both of the two target areas - dBNST and LPGi - are associated with the behavioral effects caused by the deletion of the orexin receptor 1 in dopamine neurons.
We completely agree that it would be very valuable to directly show dBNST and LPGi are associated with the behavioral effects caused by the deletion of Ox1R in dopaminergic neurons. While we are very interested in carefully investigating specific orexin-dopamine targeting areas and related neural circuits in the future, in the present manuscript, we mainly aim to give an overview of the behavioral roles of orexin-dopamine interaction and propose some promising downstream pathways.
The authors should state if data are corrected for multiple comparisons, e.g., in the PET study of different regions.
We have included information about the post-hoc tests for all 2-way ANOVA analyses in the submitted manuscript. For the PET study, the p-values in the p-maps were not corrected for multiple comparison, Figure 3—figure supplement 2 shows the raw data of each mouse and the analysis method (t-test). In the revised reviewed preprint, we include the information on the analysis method in the figure legends of Figure 3.
We consider that saline and orexin injections mimic the resting and active state of mice, respectively, and would like to study genotype effect under each condition. Doing 2-way ANOVA takes in count the difference between orexin and saline injection, which could mask the genotype effect under a certain condition. Therefore, we decided to perform t-tests for each condition in Figure 3. While we provide readers with full information in Figure 3—figure supplement 2 with the raw data of each individual mouse, below we present the p-maps after multiple comparisons (Sidak’s post hoc test). After multiple comparisons, we could see changes in similar brain regions as in Figure 3, though significant values are reduced by the correction for multiple comparisons, and under orexin-injection condition, we fail to see significantly higher activity around the lateral paragigantocellular nucleus (LPGi), nucleus of the horizontal limb of the diagonal band (HDB) and magnocellular preoptic nucleus (MCPO) in Ox1RΔDAT mice. In order to more precisely identify the anatomical locations, we performed additional experiments to confirm the changes revealed by PET. For example, LPGi is a relatively small region confirmed and identified more precisely by c-Fos immunostaining (Figure 4A, C).
Author response image 1.
PET imaging studies comparing Ox1RΔDAT and control mice, with post-hoc t-test to correct for multiple comparisons. 3D maps of p-values in PET imaging studies comparing Ox1RΔDAT and control mice, after intracerebroventricular (ICV) injection of (A) saline (NS) and (B) orexin A. Control-NS, n = 8; control-orexin, n = 6; Ox1RΔDAT, n = 8. M2, secondary motor cortex; MPA, medial preoptic area; Pir, piriform cortex; IEn, intermediate endopiriform claustrum; DEn, dorsal endopiriform claustrum; VEn, ventral endopiriform claustrum; LSS, lateral stripe of the striatum; BNST, the dorsal bed nucleus of the stria terminalis; S1Sh, primary somatosensory cortex, shoulder region; S1HL, primary somatosensory cortex, hindlimb region; S1BF, primary somatosensory cortex, barrel field; S1Tr, primary somatosensory cortex, trunk region; V1, primary visual cortex; V2L, secondary visual cortex, lateral area; SubCV, subcoeruleus nucleus, ventral part; Gi, gigantocellular reticular nucleus; IRt, intermediate reticular nucleus; sp5, spinal trigeminal tract.
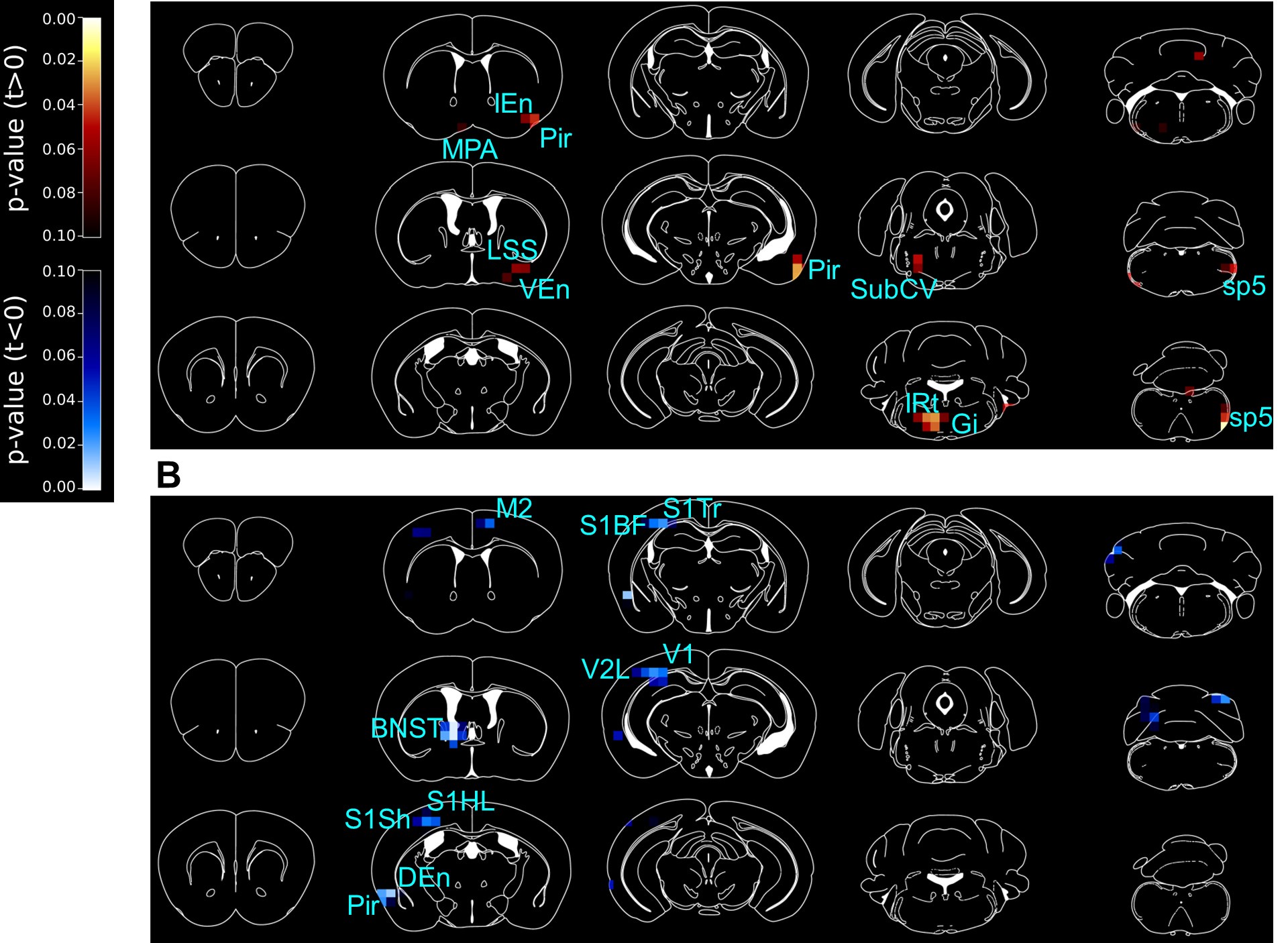
Provide a rationale for following up on BNST and LPGi and not any of the regions identified in the PET study.
We thank the reviewer for the careful reading and important input. Ox1RΔDAT mice exhibited increased novelty- and orexin-induced locomotion compared to control mice. After orexin injection, PET imaging shows that the neural activity of BNST and LPGi was lower or higher than control mice, respectively.
We selected BNST and LPGi for further validation because we think their key functional roles in regulating emotion, exploratory behaviors and locomotor speed are related to novelty-induced locomotion. We confirmed the neural activity change by c-Fos staining and investigated the expression patterns of dopamine receptors in BNST and LPGi. Our findings suggested that Ox1R deletion in dopaminergic neurons results in the disinhibition of neural activity in LPGi via dopaminergic pathways and the decrease of dopamine-mediated neural activity in BNST. Emotion perception affects the decision how to respond to the novelty. It is possible that novelty activates the orexin system and Ox1R signaling in dopaminergic neurons promotes emotion perception and inhibits exploration. Of course, further investigation is necessary to test this hypothesis in future. We have improved the rational description and discussion in the ‘Results’ and ‘Discussion’ section in the revised reviewed preprint (lines 210-213, 259-270, 293-308).
Heatmap in Fig. 1K should not have smoothing across the y-axis, individual cells should be discrete.
We thank the reviewer for bringing this issue to our attention. The data had not been intentionally smoothed (neither across the x-axis nor the y-axis), but it was probably a formatting issue. We have corrected this and separated individual cell traces with lines (Figure 1K, Figure 1—figure supplement 3).
Dopamine cells are well known to lack Fos expression in most cases. Did the authors consider using another IEG to show neural activation, e.g., pERK?
We did not use another IEG. The electrophysiological and Ca2+ imaging studies presented here, along with previous electrophysiological studies by others, clearly show that orexin A acutely and directly stimulates SN and VTA dopaminergic neurons. Please see also the response to a related comment of Reviewer 1.
Consider adding a lower magnification section to anatomical figures to aid the reader in orienting and identifying the location.
We have added the schematic illustration of SN, VTA, BNST and LPGi in Figure 1I and Figure 4— figure supplement 1. We hope this helps the reader in orienting and identifying the location.
Data availability should be stated.
There are no restrictions on data availability. We have added this section to the revised reviewed preprint.
Line 50. Some more references both historical and recent could be given to support this statement about the function of dopamine.
We have improved the description and references to support the statement about dopamine function (lines 46-58). We have cited recent studies and some reviews in the revised reviewed preprint (lines 4658).
The PET data (Fig. 3) might be easier to visualize and interpret if a white background was used. In addition, is there a more refined way of presenting the data in Fig 3, S1?
It is common to present imaging data such as PET and MRI on a black background. We also have already applied this color scheme in multiple publications and would therefore prefer to stick to this color scheme.
While Figure 3 is the concise way to present PET data, we aim to show the original individual results of mice in Figure 3—figure supplement 2 and to demonstrate how we performed the statistical analysis. Therefore, we take an example voxel of the respective brain area, perform the t-test, and present the data as bars with individual dots.
Line 97. State what type of Ca imaging here, e.g., "we performed Ca imaging in ex vivo slices of VTA and SN".
As the reviewer suggested, we have specified the type of Ca2+ imaging (line 112).
Line 165. State which groups this post-mortem analysis was performed on and if any differences were to be found (not expected to find differences in this anatomical tracing experiment but good to report this as both groups were used).
Postmortem analysis of c-Fos staining revealed low c-Fos expression in dopaminergic neurons in the VTA and SN of Ox1RΔDAT and control mice after ICV injection of saline or orexin A (1 nmol). No obvious changes were observed among the groups. We have improved the description in the revised reviewed preprint (lines 202-208).
Line 192. What do you mean by optophysiological here? The Ca imaging (which is a fairly small, confirmatory element of the manuscript).
We have changed ‘optophysiological experiments’ (line 192 in initial submitted manuscript) to ‘calcium imaging experiments’ and rephrased the beginning of the discussion to clarify the objectives (lines 238246).
The protein level in the diet is substantially higher than in most rodent diets (34% here vs 14-20% in most commercial rodent chows). Please comment on this.
This diet is for rat and mouse maintenance, purchased from ssniff Spezialdiäten GmbH (product V1554).
The percentage of calories supplied by protein is affected by the calculation methods. The company calculated with pig equation before and the value was 34% in the old instruction data sheet. They have updated the value to 23% in the new data sheet with calculations by Atwater factors. We thank the reviewer for reminding us and have updated the values in the revised reviewed preprint (lines 314-316).
Editor's note:
Should you choose to revise your manuscript, please include full statistical reporting including exact p-values wherever possible alongside the summary statistics (test statistic and df) and 95% confidence intervals. These should be reported for all key questions and not only when the p-value is less than 0.05.
We have provided the source data and the statistical reporting for each Figure with the revision
References
Baimel, C., Lau, B. K., Qiao, M., & Borgland, S. L. (2017). Projection-target-defined effects of orexin and dynorphin on VTA dopamine neurons. Cell Rep, 18(6), 1346-1355. https://doi.org/10.1016/j.celrep.2017.01.030
Korotkova, T. M., Eriksson, K. S., Haas, H. L., & Brown, R. E. (2002). Selective excitation of GABAergic neurons in the substantia nigra of the rat by orexin/hypocretin in vitro. Regul Pept, 104(1-3), 83-89. https://doi.org/10.1016/s0167-0115(01)00323-8
Korotkova, T. M., Sergeeva, O. A., Eriksson, K. S., Haas, H. L., & Brown, R. E. (2003). Excitation of ventral tegmental area dopaminergic and nondopaminergic neurons by orexins/hypocretins. J Neurosci, 23(1), 7-11. https://www.ncbi.nlm.nih.gov/pubmed/12514194
Liu, C., Xue, Y., Liu, M. F., Wang, Y., Liu, Z. R., Diao, H. L., & Chen, L. (2018). Orexins increase the firing activity of nigral dopaminergic neurons and participate in motor control in rats. J Neurochem, 147(3), 380-394. https://doi.org/10.1111/jnc.14568
Tung, L. W., Lu, G. L., Lee, Y. H., Yu, L., Lee, H. J., Leishman, E., Bradshaw, H., Hwang, L. L., Hung, M. S., Mackie, K., Zimmer, A., & Chiou, L. C. (2016). Orexins contribute to restraint stress-induced cocaine relapse by endocannabinoid-mediated disinhibition of dopaminergic neurons. Nat Commun, 7, 12199. https://doi.org/10.1038/ncomms12199
-
-
eLife assessment
This manuscript describes valuable findings on the expression pattern of orexin receptors in the midbrain and how manipulating this system influences several behaviors, such as context-induced locomotor activity and exploration. The overall strength of evidence - which includes anatomical, viral manipulation studies, and brain imaging - is solid and broadly supports claims in the paper, however, there are several areas in which the conclusions are only partially supported by the data provided. These results have implications for understanding the neural underpinnings of reward and will be of interest to neuroscientists and cognitive scientists with an interest in the neurobiology of reward.
-
Reviewer #1 (Public Review):
Summary:
In this manuscript, the role of orexin receptors in dopamine neurons is studied. Considering the importance of both orexin and dopamine signalling in the brain, with critical roles in arousal and drug seeking, this study is important to understand the anatomical and functional interaction between these two neuromodulators. This work suggests that such interaction is direct and occurs at the level of SN and VTA, via the expression of OX1R-type orexin receptors by dopaminergic neurons.Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter-expressing neurons is a strong approach to dissecting the direct role of orexin in modulating dopamine signalling in the brain. The battery of behavioural assays to study this line provides a valuable source of information for researchers …Reviewer #1 (Public Review):
Summary:
In this manuscript, the role of orexin receptors in dopamine neurons is studied. Considering the importance of both orexin and dopamine signalling in the brain, with critical roles in arousal and drug seeking, this study is important to understand the anatomical and functional interaction between these two neuromodulators. This work suggests that such interaction is direct and occurs at the level of SN and VTA, via the expression of OX1R-type orexin receptors by dopaminergic neurons.Strengths:
The use of a transgenic line that lacks OX1R in dopamine-transporter-expressing neurons is a strong approach to dissecting the direct role of orexin in modulating dopamine signalling in the brain. The battery of behavioural assays to study this line provides a valuable source of information for researchers interested in the role of orexin-A in animal physiology.Weaknesses:
The choice of methods to demonstrate the role of orexin in the activation of dopamine neurons is not justified and the quantification methods are not described with enough detail. The representation of results can be dramatically improved and the data can be statistically analysed with more appropriate methods. -
Reviewer #2 (Public Review):
Summary:
This manuscript examines the expression of orexin receptors in the midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express the orexin receptor 1 subtype and then go on to delete this receptor in dopamine neurons using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that in the absence of this receptor orexin is no longer able to excite dopamine neurons of the substantia nigra.The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and …
Reviewer #2 (Public Review):
Summary:
This manuscript examines the expression of orexin receptors in the midbrain - with a focus on dopamine neurons - and uses several fairly sophisticated manipulation techniques to explore the role of this peptide neurotransmitter in reward-related behaviors. Specifically, in situ hybridization is used to show that dopamine neurons predominantly express the orexin receptor 1 subtype and then go on to delete this receptor in dopamine neurons using a transgenic strategy. Ex vivo calcium imaging of midbrain neurons is used to show that in the absence of this receptor orexin is no longer able to excite dopamine neurons of the substantia nigra.The authors proceed to use this same model to study the effect of orexin receptor 1 deletion on a series of behavioral tests, namely, novelty-induced locomotion and exploration, anxiety-related behavior, preference for sweet solutions, cocaine-induced conditioned place preference, and energy metabolism. Of these, the most consistent effects are seen in the tests of novelty-induced locomotion and exploration in which the mice with orexin 1 receptor deletion are observed to show greater levels of exploration, relative to wild-type, when placed in a novel environment, an effect that is augmented after icv administration of orexin.
In the final part of the paper, the authors use PET imaging to compare brain-wide activity patterns in the mutant mice compared to wildtype. They find differences in several areas both under control conditions (i.e., after injection of saline) as well as after injection of orexin. They focus on changes in the dorsal bed nucleus of stria terminalis (dBNST) and the lateral paragigantocellular nucleus (LPGi) and perform analysis of the dopaminergic projections to these areas. They provide anatomical evidence that these regions are innervated by dopamine fibers from the midbrain, are activated by orexin in control, but not mutant mice, and that dopamine receptors are present. Thus, they argue these anatomical data support the hypothesis that behavioral effects of orexin receptor 1 deletion in dopamine neurons are due to changes in dopamine signaling in these areas.
Strengths:
Understanding how orexin interacts with the dopamine system is an important question and this paper contains several novel findings along these lines. Specifically:
(1) The distribution of orexin receptor subtypes in VTA and SN is explored thoroughly.
(2) Use of the genetic model that knocks out a specific orexin receptor subtype from only dopamine neurons is a useful model and helps to narrow down the behavioral significance of this interaction.
(3) PET studies showing how central administration of orexin evokes dopamine release across the brain is intriguing, especially since two key areas are pursued - BNST and LPGi - where the dopamine projection is not as well described/understood.Weaknesses:
The role of the orexin-dopamine interaction is not explored in enough detail. The manuscript presents several related findings, but the combination of anatomy and manipulation studies does not quite tell a cogent story. Ideally, one would like to see the authors focus on a specific behavioral parameter and show that one of their final target areas (dBNST or LPGi) was responsible or at least correlated with this behavioral readout. In addition, some more discussion on what the results tell us about orexin signaling to dopamine neurons under normal physiological conditions would be very useful. For example, what is the relevance of the orexin-dopamine interaction blunting novelty-induced locomotion under wildtype conditions?In some places in the Results, insufficient explanation and reporting is provided. For example, when reporting the behavioral effects of the Ox1 deletion in two bottle preference, it is stated that "[mutant] mice showed significant changes..." without stating the direction in which preference was affected.
The cocaine CPP results are difficult to interpret because it is unclear whether any of the control mice developed a CPP preference. Therefore, it is difficult to conclude that the knockout animals were unaffected by drug reward learning. Similarly, the sucrose/sucralose preference scores are also difficult to interpret because no test of preference vs. water is performed (although the data appear to show that there is a preference at least at higher concentrations, it has not been tested).
-
