Ubiquitin ligase and signalling hub MYCBP2 is required for efficient EPHB2 tyrosine kinase receptor function
Curation statements for this article:-
Curated by eLife

eLife assessment
This valuable study identifies an Ephrin type-B Receptor 2 (EPHB2) interactor, MYCBP2, as a potential regulator of EPHB2 stability and function. In contrast to expectations, based on MYCBP2 function in the ubiquitin pathway, loss of function of MYCBP2 resulted in less EPHB2 receptor and defective EPHB2 function. The paper is supported by a largely convincing set of biochemical, cell culture and in vivo experiments.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Eph receptor tyrosine kinases participate in a variety of normal and pathogenic processes during development and throughout adulthood. This versatility is likely facilitated by the ability of Eph receptors to signal through diverse cellular signalling pathways: primarily by controlling cytoskeletal dynamics, but also by regulating cellular growth, proliferation, and survival. Despite many proteins linked to these signalling pathways interacting with Eph receptors, the specific mechanisms behind such links and their coordination remain to be elucidated. In a proteomics screen for novel EPHB2 multi-effector proteins, we identified human MYC binding protein 2 (MYCBP2 or PAM or Phr1). MYCBP2 is a large signalling hub involved in diverse processes such as neuronal connectivity, synaptic growth, cell division, neuronal survival, and protein ubiquitination. Our biochemical experiments demonstrate that the formation of a complex containing EPHB2 and MYCBP2 is facilitated by FBXO45, a protein known to select substrates for MYCBP2 ubiquitin ligase activity. Formation of the MYCBP2-EPHB2 complex does not require EPHB2 tyrosine kinase activity and is destabilised by binding of ephrin-B ligands, suggesting that the MYCBP2-EPHB2 association is a prelude to EPHB2 signalling. Paradoxically, the loss of MYCBP2 results in increased ubiquitination of EPHB2 and a decrease of its protein levels suggesting that MYCBP2 stabilises EPHB2. Commensurate with this effect, our cellular experiments reveal that MYCBP2 is essential for efficient EPHB2 signalling responses in cell lines and primary neurons. Finally, our genetic studies in Caenorhabditis elegans provide in vivo evidence that the ephrin receptor VAB-1 displays genetic interactions with known MYCBP2 binding proteins. Together, our results align with the similarity of neurodevelopmental phenotypes caused by MYCBP2 and EPHB2 loss of function, and couple EPHB2 to a signalling effector that controls diverse cellular functions.
Article activity feed
-

-
-
-

Author Response
The following is the authors’ response to the previous reviews
The revised manuscript is much improved - many unclear points are now better explained. However, in our opinion, some issues could still be significantly improved.
- Statistics: none of us are experts in statistics but several things remain questionable in our opinion and if it were our study, we would consult with an expert:
a) while we understand the authors note about N-chasing and p-hacking, we wonder how the number of N's was premeditated before obtaining the results. Why in 4M an N of 3 is sufficient while in 3E the N is >20 (and not mentioned). At the very least, we think it would be wise to be cautious when stating something as not-significant when it is clear (as in 4M) that the likelihood of it actually being statistically significant is quite …
Author Response
The following is the authors’ response to the previous reviews
The revised manuscript is much improved - many unclear points are now better explained. However, in our opinion, some issues could still be significantly improved.
- Statistics: none of us are experts in statistics but several things remain questionable in our opinion and if it were our study, we would consult with an expert:
a) while we understand the authors note about N-chasing and p-hacking, we wonder how the number of N's was premeditated before obtaining the results. Why in 4M an N of 3 is sufficient while in 3E the N is >20 (and not mentioned). At the very least, we think it would be wise to be cautious when stating something as not-significant when it is clear (as in 4M) that the likelihood of it actually being statistically significant is quite large.
b) In most analyses, the data is not only normalized by actin or some other measure but also to the first (i.e left side on the graph) condition, resulting in identical data points that equal '1' (in Figure 4 alone - C; I; K; M; and O) - while this might be scientifically sound, it should be mentioned (the specific normalization) and also note that this technique shadows any real variance that exists in the original data in this condition. consider exploring techniques to overcome this issue.
c) In 3C, - if we understand the experiment, you want to convince us that the DIFFERENCE between eB2-FC compared to FC is larger in the control compared to the experiment. We are not absolutely sure that the statistical tools employed here are sufficient - which is why we would consult an expert.
A) We are aware that many studies do not consistently quantify such experiments. For example, there are essentially no published examples of the signalling timelines of EphB2 receptors as in Fig. 5. By striving to quantifying such biochemical effects, an unquantified experiment stands out, and so perhaps we were too strict by trying to quantify as many experiments as possible, resulting in low n’s for some of them. We acknowledge that additional experiments on EPHB1 protein stability may reach significance. We have adjusted our text on line 332-335 to point to this interesting trend, and slightly changed the conclusion to this section. Similarly, we commented on similar trends when describing Figs. 1E and 4G on lines 901 and 952.
B) For the Western blot band intensity normalisation, we believe that our method is scientifically sound. Normally, when the replicate samples are loaded on one gel and blotted on the same membrane, the experimenter only needs to normalise the target band intensity to its cognate loading control band intensity for quantitation. However, we usually have a large number of samples from multiple experiments, carried out on different dates. For example, in Fig. 4B,C there are 7 biological replicates collected from 7 experiments and in Fig. 4D there are 10 protein samples. It is not possible for us to run all samples on the same gel. In addition, due to the combined effects of variance in transfer efficiency, the potency of antibodies, detection efficiency and the developing time for each blot, it is practically impossible to generate similar band intensity for each batch. Thus, we use normalisation of test bands to the loading control for individual experiments, and this analysis method is widely accepted by reputable journals with a focus on biochemical experiments (for example: PMID 37695914: Fig. 3 A,B,C; PMID 36282215: Fig. 3 B,C,D,E; PMID 33843588: Fig. 3 C,D,E,F,G,H). Since the value of the first sample on the plot is 1, which is a hypothetical value and does not meet the parametric test requirement, we performed one-sample t-test for statistics when other samples are compared with the first sample (PMID 35243233 Fig. 6 A,B,C,D; https://www.graphpad.com/quickcalcs/oneSampleT1/, “A one sample t-test compares the mean with a hypothetical value. In most cases, the hypothetical value comes from theory. For example, if you express your data as 'percent of control', you can test whether the average differs significantly from 100.”). Thus, we believe that our normalisation and statistical methods are both correct with a large number of precedents.
C) This comment refers to the cell collapse experiment shown in Fig. 3C for which the data are plotted in Fig. 3D. We stand by the statistical method used. There are two groups of cells (CTRLCRISPR and MYCBP2 CRISPR) and two treatments for each cell group (Fc control and eB2), thus we should use two-way ANOVA. Since we compared the cell retraction effects of Fc and eB2 on the two groups of cells, Sidak post hoc comparison is the right method to avoid errors introduced by multiple comparisons. Here is an example of an eLife article that used the same statistical method for similar comparisons: PMID 37830910, Fig. 1 H,I. To make the comparison easier, we grouped the experiments by cell type (CTRLCRISPR and MYCBP2 CRISPR) as opposed to by treatment. Below, the old version is on the right, and the new version is on the left. The conclusion is that eB2 induces less cell collapse in cells depleted of MYCBP2, when compared to the control cells. However, eB2 is still able to collapse cells lacking MYCBP2.
Author response image 1.
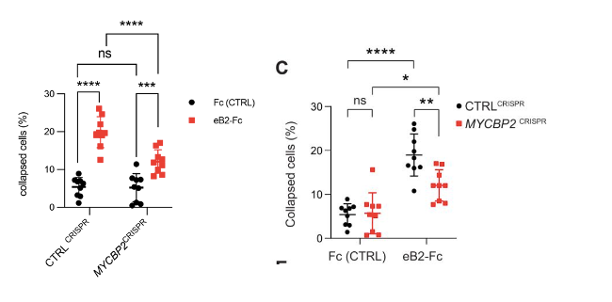
Revisiting these data, we noticed an error introduced when CC compiled the data used to generate Fig. 3D. The data were acquired from nine biological replicates per condition. CC used a mix of two methods for cell collapse rate calculation: the first method involved the sum of collapsed cells and all cells from multiple regions of one coverslip (biological replicate). The second method involved computing a collapse rate in each region which then was used to calculate the average collapse rate for the entire coverslip (technical replicate). Given the small cell numbers due to sparse culture conditions, we believe that the first method is a more conservative approach. We hence re-plotted all replicate data using the first method. This resulted in slightly different % collapse and p values. These were changed accordingly in the text and plot and do not affect the conclusion of this experiment.
- thanks for the clarification that the interaction between the extracellular domain of EPHB2 and MYCBP2 might not occur directly - however, unless we missed this it was not clearly stated in the text. It is an important point and also a cool direction for the future - to find the elusive co-receptor that actually helps EPHB2 and MYCBP2 form a complex.
We now also refer to this in the results section on line 215.
“Since EPHB2 is a transmembrane protein and MYCBP2 is localised in the cytosol, these experiments suggest that the interaction between the extracellular domain of EPHB2 and MYCBP2 might be indirect and mediated by other unknown transmembrane proteins.”
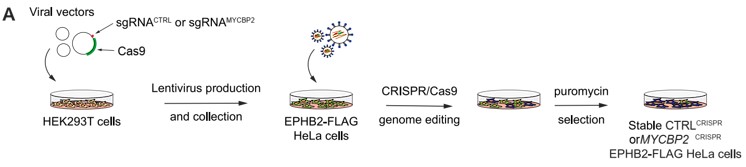
- The Hela CRISPR cell line is better explained in the response letter but still not sufficiently explained in the text for a non-expert reader. If the authors want any reader to comprehend this, we would strongly recommend adding a scheme.
We now include a schematic outlining the CRISPR cell generation as Fig. 3A and its description on line 926.
Author response image 2.
- To clarify some of our previous (and persisting) concerns about Figure 3D/E - it is true that a reduction in 25% of cell size is dramatic. But (if we understand correctly) your claim is that a reduction in 22% (this is a guess, as the actual numbers are not supplies) is significantly less than 25%. Even if it is, statistically speaking, significant, what is the physiological relevance of this very slight effect? In this experiment, the N was quite large, and we wonder if the images in D are representative - it would be nice to label the data points in E to highlight which images you used.
We now mention the average cell area contraction measurements in the legend to Fig. 3F on line 935. We also tracked down the individual cells shown in Fig. 3E and they are now labelled as data points in blue in Fig. 3F. HeLa cell collapse is a simplified model of EPHB2 function and we do not know whether the difference between the behaviour of CTRLCRISPR and MYCBP2 CRISPR cells is physiologically significant and thus we prefer not to speculate on this.
- Figure 3F and other stripe assays - In the end, it is your choice how to quantify. We believe that quantifying area of overlap is a more informative and objective measurement that might actually benefit your analyses. That said, if you do keep the quantification as it is now, you have to define the threshold of what you mean by "cell/s (or an axon in 7A, where it is even more complicated as are you eluding to primary, secondary, or even smaller branches) are RESIDING within the stripe". Is 1% overlap sufficient or do you need 10 or 50% overlap?
We now added this statement to the methods on line 745: “A cell was considered to be on an ephrin-B2 stripe when more than 50% of its nucleus was located on that stripe”. For chick explant stripe assay, when measuring the length of an axon on a stripe, we only measured the main axons originated from the explants.
For explant/stripe experiments in Fig. 7 AB, we now use the term “GFP-expressing neurite” rather than “branch”. This was already present in the results of the previous version, but the methods and legend needed to be brought up to date (lines 786 and 1008. We think that “branch” was a confusing term that was supposed to mean the same thing as “neurite” but came across as some indication of branching. We do not know whether the GFP+ neurites were primary or secondary extensions of explants, or in fact, whether some of them contained more than one axon. We also adjusted the method to reflect the fact that some stripes were used in conjunction with a single explant and added a reference to a previous study extensively using this method (Poliak et al., 2015) on line 778.
- We still don't get the link to the lysosomal degradation. Your data suggests that in your cells EPHB2 is primarily degraded by the lysosomal pathway and not proteasome. Any statement about MYCBP2 is not strongly supported by the data, in our opinion - Unless you develop some statistical measurement that shows that the effect of BafA1 is statistically different in MYCBP2 cells than in control cells. Currently, this is not the case and the link is therefore not warranted in our opinion.
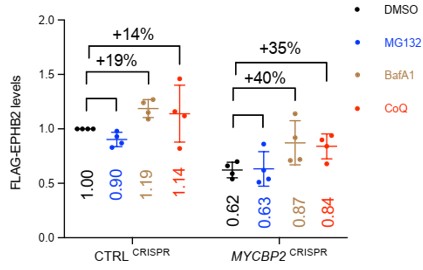
We generated a new version of Fig. 4K with average increase in EPHB2 levels in the presence of BafA1 and CoQ, compared to DMSO treated controls (see below). BafA1 and CoQ restored EPHB2 protein levels by 19% and 14% respectively in CtrlCRISPR cells, while the inhibitors restored EPHB2 protein levels by 40% and 35% respectively in MYCBP2 CRISPR cells.
Author response image 3.
For each of the 4 replicates, the increase in EPHB2 levels by BafA1 compared to DMSO is as follows:
Author response table 1.
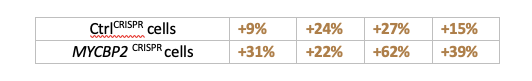
These values are not significantly different between CtrlCRISPR cells versus MYCBP2 CRISPR cells (p= 0.08, student’s t test). Similarly for the CoQ experiment. We now temper our conclusion for this experiment: Although the difference in percentage increase between CTRLCRISPR cells and MYCBP2CRISPR cells is not significant, this trend raises the possibility that the loss of MYCBP2 promotes EPHB2 receptor degradation through the lysosomal pathway (line 319). We also adjusted the section title (line 306).
- While the C. elegans part is now MUCH better explained - we are not sure we understand the additional insight. The fact that vab-1 and glo4 double mutants are additive as are vab1 and fsn1, suggest they act in parallel (if the mutants are NULL, and not if they are hypomorphs, if one wants to be accurate) - how this relates to your story is unclear. The vab1/rpm1 double mutant is still uninformative and incomplete. rpm1 phenotype is so severe that nothing would make it more severe. We read the Jin paper that the authors directed to - nothing makes the rpm1 phenotype more severe. Yes, some DOWNSTREAM elements make the rpm1 phenotype LESS severe - this is not something you were testing, to the best of our knowledge. Rather, you wanted to see if rpm1 mutant resulted in stabilization of vab1 and thus suppression of vab1 phenotype - we are just not sure the system is amenable to test (actually reject) your hypothesis that Vab1 is degraded by rpm1. Also, assuming we are talking about NULLs, the fact that the rpm1 phenotype is WAY stronger than the vab1 mutant, suggests that rpm1 functions via multiple routes, adding even more complexity to the system. Given these results, despite the much improved clarity, we are still not sure that the worm data adds new insight, rather than potentially confusing the reader.
We realise that the genetic interactions between vab-1 and the RPM-1/MYCBP2 signalling network are complicated. However, we insist on keeping the data for the sake of its availability for future studies and completeness. We also think it is important for readers and the community to see these data, even if the authors and reviewers are not entirely in agreement about the importance/interpretation of experimental outcomes. It is our hope that the community will examine the results and draw their own conclusions.
A few points of clarification:
The C. elegans experiments were designed to test genetically if the vertebrate interactions between EPHB2 and MYCBP2 and its signalling network are conserved. We studied two kinds of interactions: (1) between vab-1 and RPM-1/MYCBP2 downstream proteins (GLO-4 and FSN-1) and (2) between vab-1 and rpm-1. For these studies, we used null alleles for vab-1, glo-4 and fsn-1 which is now noted on lines 440, 453, 475 and 859. Our findings are consistent with the VAB-1 Ephrin receptor functioning in parallel to known RPM-1 binding proteins. This is further supported by new data: vab-1; fsn-1 double mutants showed enhanced incidence of axon overextension defects using a second transgenic background, zdIs5 (Pmec-4::GFP), to visualize axon termination (Fig. 8F).
This second transgenic background also allowed us to generate new data to address your concerns about phenotypic saturation in rpm-1 mutants. To do this, we used the zdIs5 (Pmec4::GFP) genetic background, in which axon termination defects are not saturated in rpm-1 mutants (Fig. 8F) because they can be enhanced by other mutants such as cdc-42 and unc-33 (Fig. 7C, D, in Borgen et al. Development 144, 4658–4672 (2017), PMID 29084805). In this new background, we found that vab-1 loss of function fails to enhance the incidence of severe “hook” defects in rpm-1 mutants which is an indication that the two genes function in the same pathway. Importantly, prior studies in this background, also showed that mutants in the RPM-1 signalling network (e.g. fsn-1, glo-4 and ppm-2) do not enhance the incidence of severe “hook” defects as double mutants with rpm-1 compared to rpm-1 single mutants (Fig. 7B, ibid.).
To reflect these ideas more clearly, we revised the Results section pertaining to C. elegans genetics (starting on line 418) and tempered our discussion (lines 517). Basically, this section now says that we studied genetic interactions between vab-1 and the RPM-1/MYCBP2 signalling network. From these experiments we conclude that: (1) The enhancement of overextension defects in vab-1; glo-4 and vab-1; fsn-1 double mutants compared to single mutants indicates that VAB-1/EPHR functions in parallel to known RPM-1 binding proteins to facilitate axon termination, and (2) Since the vab-1; rpm-1 double mutants do not display an increased frequency or severity of overextension defects compared to rpm-1 single mutants, VAB-1 /EPHR functions in the same genetic pathway as RPM-1/MYCBP2.
The new genetic data included in this version were generated by Karla J. Opperman who is now included as a co-author.
Further corrections:
Author response image 4.
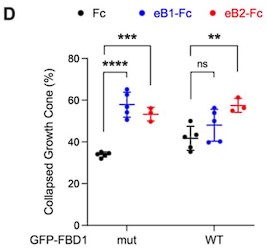
Because of the errors associated with quantifications in Fig. 3D (see above), we reviewed other quantification methodologies and noticed another discrepancy that required a correction. In the hippocampal neuron growth cone collapse assay shown in the previous version of Fig. 7 D (left), the growth cones were classified into three groups: 1, fully collapsed; 2, hard to tell, but not fully collapsed; 3, fan-shape cones. Two different quantifications were performed as follows: (1), number of fully collapsed cones divided by the numbers of all growth cones; (2), number of fully collapsed cones divided by [number of fully collapsed cones + fan-shape cones]. CC erroneously used the second method to generate Fig. 7D.
We think that the first method is more appropriate. Furthermore, since n=5 for the Fc and eB1-Fc conditions, but n=3 for the eB2-Fc condition, we decided to omit it. The final plot for figure 7D is the following:
Author response image 5.
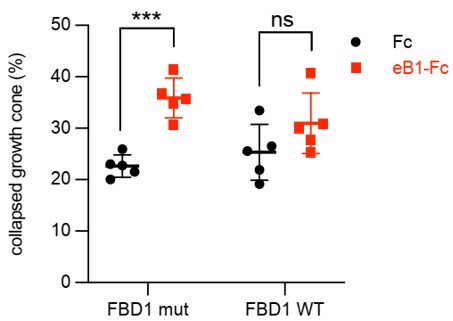
Our conclusion still stands that exogenous FBD1 WT overexpression impaired the growth cone collapse mediated by EphB.
-
eLife assessment
This valuable study identifies an Ephrin type-B Receptor 2 (EPHB2) interactor, MYCBP2, as a potential regulator of EPHB2 stability and function. In contrast to expectations, based on MYCBP2 function in the ubiquitin pathway, loss of function of MYCBP2 resulted in less EPHB2 receptor and defective EPHB2 function. The paper is supported by a largely convincing set of biochemical, cell culture and in vivo experiments.
-
Reviewer #1 (Public Review):
The Eph receptor tyrosine kinase family plays a critical function in multiple physiological and pathophysiological processes. Hence, understating the regulation of these receptors is a highly important question. Through extensive experiments in cell lines and cultured neurons Chang et.al show that the signaling hub protein, MYCBP2 positively regulates the overall stability of a specific member of the family, EPHB2, and by that the cellular response to ephrinBs.
Overall, this work sheds light on the divergent in the regulatory mechanisms of the Eph receptors family. Although the physiological importance of this new regularly mechanism in mammals awaits to be discovered, the authors provide genetic evidence using C.elegans that it is evolutionarily conserved. -
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) is also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed …
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) is also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed that FBXO45 within a ternary complex increased the stability of the EphB2/MYCBP2 complex.
To define the interacting domains, they used clearly designed swapping experiments to show that the extracellular and transmembrane domains are necessary and sufficient for the formation of the ternary complex.
Using a cellular contraction assay, the authors showed the necessity of MYCBP2 in mediating the cytoskeletal response of EphB2 forward signalling. Furthermore, they used the technically challenging stripe assay of alternating lanes of ephrinB-Fc and Fc to show that also in this migration-based essay MYCBP2 is required for EphB mediated differential migration pattern.
MYCBP2 in addition is necessary to stabilize EphB2, that is in the absence of MYCBP2, EphB2 is degraded in the lysosomal pathway.
Interestingly, the third protein in this complex, Fbxo45, was further characterized by overexpression of the domain of MYCBP2, known to interact with Fbxo45. Here the authors showed that this approach led to the disruption of the EphB2 / MYCBP2 complex, and also abolished the ephrinB mediated activation of EphB2 receptors and their differential outgrowth on ephrinB2-Fc / Fc stripes.
Finally, the authors demonstrated an in vivo function of this complex using another model system, C elegans where they were able to show a genetic interaction.
Data show in a nice set of experiments a novel level of EphB2 forward signalling where a ternary complex of this receptor with multifunctional MYCBP2 and Fbxo45 controls the activity of EphB2, allowing a further complex regulation of this important receptors. Additionally, the authors challenge pre-existing concepts of the function of MYCBP2 which might open up novel ways to think about this protein.
Of interest is this work also in terms of development of the retinotectal projection in zebrafish where MYCBP2/highwire plays a crucial role, and thus might lead to a better understanding of patterning along the DV axis, for which it is known that EphB family members are crucial.Overall, the experiments are classical experiments of co-immunoprecipitations, swapping experiments, collapse assays, and stripe assays which all are well carried out and are convincing.
-
Reviewer #3 (Public Review):
In this improved version of the manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, suggesting the involvement of additional proteins as MYCBP2 is thought to be a cytoplasmic protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 …
Reviewer #3 (Public Review):
In this improved version of the manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, suggesting the involvement of additional proteins as MYCBP2 is thought to be a cytoplasmic protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 as a ubiquitin E3 ligase, it actually positively regulates EPHB2 protein stability, and function.
The strength of this manuscript is the extensive biochemical analysis of the EPHB2/MYCBP2/FBXO43 interactions. The vast majority of the conclusions are supported by the data.
The attempt to extend the study to an in vivo animal using the worm is important, however the additive insight is, unfortunately, minimal.
-
-
Author Response
The following is the authors’ response to the original reviews.
We thank the reviewers for their thoughtful assessment of our work and their valuable critiques which we will address in the “Recommendations for the authors” section below. In particular, we appreciate Reviewer #3 noting the value of the C. elegans model system and our efforts to bridge models with our study. We agree with the reviewer that there is a need to clarify the rationale, presentation and interpretation of our results. We have substantially revised the text in our manuscript and Figure legend to address this issue, and provided extensive new commentary and citations to lay out the logic behind our experiments. Indeed, it was our oversight not being more thorough about this initially. We have further adjusted our conclusions to be less unequivocal. …
Author Response
The following is the authors’ response to the original reviews.
We thank the reviewers for their thoughtful assessment of our work and their valuable critiques which we will address in the “Recommendations for the authors” section below. In particular, we appreciate Reviewer #3 noting the value of the C. elegans model system and our efforts to bridge models with our study. We agree with the reviewer that there is a need to clarify the rationale, presentation and interpretation of our results. We have substantially revised the text in our manuscript and Figure legend to address this issue, and provided extensive new commentary and citations to lay out the logic behind our experiments. Indeed, it was our oversight not being more thorough about this initially. We have further adjusted our conclusions to be less unequivocal. Finally, we added an RPM-1 signaling diagram (Fig. 8A) to more clearly annotate the players in the RPM-1/MYCBP2 signaling network that were evaluated genetically in Fig. 8. Importantly, we provide clearer commentary on how genetic enhancer effects with known RPM-1 binding proteins and the absence of genetic suppression in vab-1/Eph receptor double mutants with components of the RPM-1/FSN-1 ubiquitin ligase complex are consistent with the biochemical finding that MYCBP2 stabilizes but does not degrade EphB2. Text edits reflecting these points are in the abstract, the C. elegans results section starting on line 411, and the discussion on lines 499, 502-504 and 541.
Following extensive discussions between the three reviewers, all three agree that the C. elegans data, as presented, does not add to, and in fact might harm, your bottom line. Our combined suggestion is to take this data out unless you plan to improve it substantially. All reviewers are perplexed by Figure 2F and the presumed interactions of cytosolic proteins with the extracellular domain of EPHB2. At the very least, please provide some suggestions/model/interpretation.
We have adjusted our manuscript substantially to address this. Please see detailed comments in the individual Reviewer sections below.
We would like to thank the reviewers for their thorough examination of our manuscript, constructive criticisms, and helpful suggestions.
Reviewer #1 (Recommendations For The Authors):
The work is extensive in my view, and mostly of high quality. See minor comments on some of the figures below.
Thank you very much.
Two more major comments :
- I don't think the C. elegans work adds to - in fact I think it hurts - the statement that this regulatory mechanism is specific to EphB2. I would advise the authors to take it out.
We agree that C. elegans has a sole Eph receptor called VAB-1 and is therefore not a specific model for EPH2B. However, testing MYCBP2 specificity for EPHB2 was not the goal or our perceived value for the C. elegans experiments. We now clarify this in the text of the Results section.
Rather, we are providing evidence that the C. elegans ephrin receptor interacts genetically with known MYCBP2/RPM-1 binding proteins. Moreover, we now provide an extensive array of citations to note that genetic enhancer interactions between different RPM-1/MYCBP2 binding proteins is well established. The reviewer has nicely highlighted for us that we handled the C. elegans genetics in too cursory a fashion in our original manuscript. We appreciate this being noted and have now aimed to make this substantially clearer. We hope the reviewer agrees that our revised C. elegans section accomplishes this goal.
Furthermore, we extensively revised the text of the Results to emphasize a key point: our observation that axon termination defects are not suppressed in vab-1; fsn-1 and vab-1; rpm-1 double mutants excludes the possibility that the VAB-1 Eph receptor is a substrate that is inhibited or degraded by the RPM-1/FSN-1 ubiquitin ligase complex. If the VAB-1 Eph receptor were ubiquitinated and degraded by the RPM-1/FSN-1 complex, we would have observed a suppression of phenotype in vab-1; rpm-1 double mutants. The precedent for this genetic relationship between the RPM-1 ubiquitin ligase and its substrates that are degraded has been established by several prior studies (PMID: 15707898; PMID: 31676756; PMID: 35421092). We now more clearly note that the absence of genetic suppression in vab-1; rpm-1 double mutants and vab-1; fsn-1 double mutants is consistent with the non-canonical stabilizing role of MYCBP2 on EPHB2 that was observed in our biochemical experiments with mammalian cells.
We also adjusted the text of the manuscript to stress that we are testing genetic interactions between the VAB-1 Eph receptor and known RPM-1 binding proteins. This is a key point, as genetic enhancer interactions are consistent with the Eph receptor functioning in the RPM-1 signaling network. This concept has been well established for RPM-1 binding proteins as now noted in our revised text with an extensive number of additional citations to published work.
Based on the above arguments, we respectfully disagree with the reviewer that our C. elegans data should be removed from the paper. To re-iterate, we are not trying to evaluate specificity for MYCBP2 and EPHB2 in C. elegans. Rather, our goals are twofold: 1) To ask whether there is an evolutionarily conserved functional genetic link between Eph receptors and known RPM-1 binding proteins. 2) To provide further in vivo genetic evidence invalidating the hypothesis that Ephrin receptors could be ubiquitination substrates that are inhibited/degraded by MYCBP2.
Text edits reflecting these points are in the abstract, the C. elegans results section starting on line 411, and the discussion on lines 499, 502-504 and 541.
- The cellular responses are not robust and the effects of MYCBP2 KO - although significant - are minor in most cases. But I don't think more experiments will help here.
We interpret the comment about the robustness to mean that the extent to which a given cellular response is affected by the loss of MYCBP2 is minor. First, the cellular responses themselves are typical of previous studies and depend on the cellular biology underlying them. For example, a growth collapse of ~50-60% over a background of 10% (Fig. 7) is typical for these sorts of assays (PMID: 37369692; PMID: 33972524; PMID: 17785182). A decrease of cell area by ~25% (Fig. 3) is quite substantial if one considers how much of a cell’s volume is taken up by the nucleus and organelles. Second, the phenotypes elicited by the loss of MYCBP2 are likely brought on by a decrease in EphB2 protein levels, but not its complete absence, as suggested by our biochemical experiment. Given that EphB2 complete loss only affects the cellular responses to a limited extent, the minor effects are not a surprise (e.g. for GC collapse: PMID: 23143520). Nevertheless, the subtle changes in cellular phenotypes, elicited by EPHB2 signaling are often sufficient to achieve proper cell positioning and cell response to guidance cues. For instance, regulation of the growth cone collapse of the outgrowing axons requires delicate changes that are dynamic and temporal.
Minor:
Fig 1C - EPHA3 and EPHB2 seem to run in different sizes, is this the case? In 2A they run at the same size.
We believe this size discrepancy is due to different percentages of SDS-PAGE gels used to resolve proteins. In Fig. 1C, we used a 6% gel for a Western blot analysis of both EPHA3/-B2-FLAG (~130 kDa) and MYCBP2 (~510 kDa). In Fig. 2A however, we performed Western blot analysis using 10% resolving gel to separate and detect EPHA3/-B2-FLAG along with MYC-FBXO45 (~30 kDa). We have reviewed the results obtained from additional biological replicates of this experiment, and observed a similar pattern in gel migration of EPHA3/-B2-FLAG across all replicates.
Fig1F - I can't trust the MYCBP2 blot.
Indeed, the MYCBP2-EPHB2 co-IP with endogenous proteins was not convincing. We now repeated this experiment using rat cortical neurons, and the results replace the previous Fig. 1F panel as mentioned on line 158.
In Fig2b the authors claim that there is enhancement in the binding of MYCBP2 and EPHB2 upon FBXO45 expression. For this type of statement quantification is required.
The quantification is now included in Fig. 2C and its significance is mentioned on line 180. Our conclusion about the enhancement stands.
Fig2G - it remained unclear to me where the binding site to MYCBP2 is, how long is the cytoplasmic tail in the DeltaICD protein?
Based on our experimental observations from Fig. 2E-H, we concluded that the fragment encompassing the extracellular domain(s) and/or transmembrane (TM) domain of EPHB2 is necessary for the protein complex formation with MYCBP2. We would like to accentuate that the EPHB2-MYCBP2 interaction might not be direct, and might involve other transmembrane protein(s) acting as a scaffold for EPHB2 and MYCBP2 binding. We did not pursue experiments to determine the exact region of the extracellular-TM portion of EPHB2 that is required for the interaction with MYCBP2.
The cytoplasmic tail in ΔICD protein consists of 25 aa of the N-terminal fragment of EPHB2 juxtamembrane (JM) region, which is adjacent to the TM helix, and followed by the 8 aa FLAG tag (EPHB2 ΔICD domain composition: extracellular domains – TM domain – 25 aa fragment of JM region – FLAG). We have determined the TM and JM sequences based on Hedger et al. (PMID: 25779975) and included the N-terminal portion of the JM region to facilitate proper ΔICD protein localization within the plasma membrane (PMID: 35793621). We modified the schematic in Fig. 2G to better visualise the EPHB2 truncations and now provide information on their size in the figure legend.
Always good to have a model of how all these proteins work together.
While we acknowledge that this would be helpful, we do not have a clear answer on how the EPHB2-MYCBP2 complex formation occurs. This requires further elucidation of the putative proteins involved in this ternary complex or testing the possibility that a MYCBP2 fragment is extruded extracellularly. Without these experiments there are too many possibilities to summarise into a clear model figure. We thus did not make any edits regarding these possibilities in the section starting on line 195.
Reviewer #2 (Recommendations For The Authors):
Overall, the experiments are classical experiments of co-immunoprecipitations, swapping experiments, collapse assays, and stripe assays which all are well carried out and are convincing.
Thank you for your encouraging comments.
Controls for the stripe assay may include Fc / Fc stripe assays.
We have performed these control experiments and now include their quantifications in the results sectioning concerning Fig. 3, starting on line 249, and those concerning Fig. 6 on line 381.
It is not clear to me why SD and not SEM has been used here for presentations.
Standard deviation (SD) measures the dispersion of a dataset relative to its mean. The standard error of the mean (SEM) measures how much discrepancy is likely in a sample’s mean compared with the population mean. Thus, SEM includes a statistical inference about the sampling distribution while SD is a less “processed” measurement that by definition is larger than SEM. SEM might make the data look less dispersed and many journals encourage the use of SD in bar graphs (PMID: 16223828).
Fig 7A: it is rather difficult to see 'branches' in Fig. 7A, better pictures and close-ups should be provided. How are branches defined? This piece of work needs more attention.
To remedy this shortcoming, we now provide inverted images with GFP signal in dark pixels overlaid on Fc (white) / eB2 (pink) stripes next to the original images.
Reviewer #3 (Recommendations For The Authors):
- My most important suggestion to the authors would be to more carefully describe the results and their interpretation of the results. Sometimes, the distinction is not clear.
We modified the text throughout the manuscript to address this.
- There are several cases, when the authors report on trends that are not statistically significant (1D, for example), or report no change, when it is clear that the addition of one more sample could have dramatically made a difference (4M - see point 12).
We agree that some of the nonsignificant differences could become significant if we added more Ns. But we prefer not to move our experimental design towards N-chasing and p-hacking (PMID: 25768323). The number of biological replicates is normally pre-determined before the onset of the experiment. Of course, some replicates can be discarded if there is a valid reason, such as a technical issue with the experiment or a positive control not working but this is not relevant for the dataset we have provided.
- Data in 1F is very difficult to interpret.
As in response to Reviewer #1: Indeed, the MYCBP2-EPHB2 co-IP with endogenous proteins was not convincing. We now repeated this experiment using rat cortical neurons, and the improved results are in revised Fig. 1F.
- Figure 2 puts Figure 1 in a strange perspective. If I understand correctly, fig 2 claims that EPHB2 interaction with MYCBP2 depends on FBXO45 - if that is the case then how does the binding in Figure 1 occur?
Indeed, we propose that the EPHB2-MYCBP2 interaction depends on FBXO45. In Fig. 2, we reveal that FBXO45 enhances the formation of the EPHB2-MYCBP2 complex. Thus, we suspect that the endogenous FBXO45 present in HeLa cells and neurons would mediate the interaction between EPHB2 and MYCBP2 in Fig. 1 experiments. We were unable to show this by Western blotting due to lack of reliable commercial antibodies against FBXO45, the complex containing endogenous FBXO45 and EPHB2 is also implied by our AP-MS data (Fig. 1B) and published databases.
- I am still trying to wrap my mind around the results in 2G-H. So do MYCBP2 and FBXO45 bind the extracellular domain of EPHBP2? What does that mean?
(see also our response to Reviewer #1, end of their section) Based on our experimental observations from Fig. 2G-H, we conclude that the fragment encompassing the extracellular domain(s) and/or transmembrane domain of EPHB2 is necessary for the protein complex formation with MYCBP2 and FBXO45. Although there is a possibility that MYCBP2 directly binds the extracellular portion of EPHB2, we have not formally tested this hypothesis. MYCBP2 has been previously shown to interact with the extracellular portion of transmembrane N-cadherin (CDH2) via BioID proximity labeling and AP-MS proteomics approaches (PMID: 32341084).
Considering the results in Fig. 2A-B, we suspect that EPHB2-MYCBP2 interaction is indirect, as FBXO45 enhances this association. Secretion of FBXO45 and direct binding of FBXO45 to the extracellular cadherin (EC1-2) domains of N-cadherin has been documented (PMID: 25143387; PMID: 32341084). Although, not tested, this is also a possibility for EPHB2-FBXO45 mode of interaction. Nevertheless, we also cannot rule out the possibility that an unknown transmembrane protein binds EPHB2 extracellularly and the same unknown protein binds MYCBP2/FBXO45 intracellularly. Resolving this model is beyond the scope of this study and will require us to pursue extensive new lines of investigation.
- I don't understand the stable Hela cell line CRISPR - is this a stable MYCBP2 deletion? In which case why is there only a reduction, not complete elimination of the protein? Or, is this a stable integration of a plasmid generating gRNA against MYCBP2? In which case, I would expect a homozygous null to emerge at some point. In any case, this is not well explained.
These lines are not derived from single cells infected with the CRISPR sgRNA-carrying viruses, therefore they are not clonal and probably contain some cells that express normal levels of MYCBP2, hence its detection on a Western. This is now clarified starting on line 221 and on line 608.
- In 3C - is this the right statistical analysis?? I would say you want to claim the different effect of the control +/- eB2 compared to the effect in the mutant +/- eB2. Still should be significant but I think a more correct analysis.
We now include this comparison in Fig. 3C as well in the results section starting on line 234.
- The robustness of the assay in Figure 3D is underwhelming – how was the area measured?
This is a live imaging experiment. Fig. 3D plots cell area at 60 minutes after ephrin-B2 addition as a fraction of the same cell’s area at 0 minutes (ephrin-B2 addition). For control cells that is a decrease of ~25%. If one considers that a cell’s nucleus and organelles like the Golgi Apparatus take up most of its volume, the magnitude is not that surprising.
- Figure 3F – did you try to plot the relative area of overlap divided by the total cellular area? You might get a more striking phenotype. Also – claiming that this confirms that MYCBP2 is REQUIRED for EPHB2 function is a bit overstated, especially given that we don’t know (do you?) the EPHB2 mutant phenotype in this assay.
We preferred to stay with the original method of image quantification which we use for other assays. With respect to the requirement of MYCBP2 for EPHB2 function in the stripe assay, our logic is rooted in the observation that native HeLa cells do not respond to ephrin-B2 stripes (45.46 ± 7.62% of cells on eB2 stripes v. Fc; data not shown). When they are transfected with EPHB2 expression plasmids they do, therefore we assume that EPHB2 expression endows them with a sensitivity to eB2 stripes. A loss of MYCBP2 attenuates this sensitivity. We clarified this starting on line 246 and on line 251.
- I didn't quite get the difference between 4A and 4B.
We apologize for the confusion. In Fig 4A, we used a stable HeLa cell line that has tetracycline-inducible expression of EPHB2-FLAG. Using these cells, we subsequently generated CTRLCRISPR or MYCBP2CRISPR cells. In these cells we then induced EPHB2 expression with tetracycline and observed that deletion of MYCBP2 resulted in the reduction of EPHB2 protein levels. To confirm this observation and to rule out the possibility that EPHB2 protein reduction is an effect of the CRISPR lines generation, we tested whereas MYCBP2 deletion reduces EPHB2, which has been transiently overexpressed (Fig. 4B). We hence conclude that loss of MYCBP2 decreases EPHB2 that was either expressed from a stable locus (Fig. 4A) or from transient transfection (Fig. 4B). We modified the Results section starting on line 262 to make this point clear.
- The entire link to lysosomal degradation should be strengthened. Perhaps I am confused, but if the reduced EPHB2 levels in MYCBP2 mutant cells result from impaired lysosomal degradation then inhibiting the lys-deg should bring the protein levels back to normal (i.e. CRISPR control) - no? As currently presented, I do not understand nor do I think the claim is strongly supported by the data.
Before treatment with inhibitors, EPHB2 levels in MYCBP2CRISPR cells are already 40% lower than they are in CTRLCRISPR cells and in all our attempts, inhibitors can only rescue/restore EPHB2 in MYCBP2CRISPR cells to a level that is lower than in CTRLCRISPR cells. But this restoration is greater in MYCBP2CRISPR than in MYCBP2CTRL cells (BafA1: 19% increase in CTRL cells and 40% in MYCBP2CRISPR cells; CoQ: 10% comparing to 35%). This indicates that EPHB2 degradation through the lysosomal pathway in MYCBP2CRISPR cells is stronger, explaining why EPHB2 degradation is promoted in MYCBP2CRISPR cells, compatible with reduced EPHB2 levels and enhanced EPHB2 ubiquitination.
- 4M, O - reporting ns based on these data seems a bit strange to me... Add one point and it will be strongly significant.
See our response to point (2), above. We prefer not to invoke potential p-hacking.
- 7d - so what are you claiming? That the cellular response to eB1 but not eB2 is affected by the addition of FBD1? this is almost the opposite of what you wrote in the text...
We treated the cells with two different ephrin-B ligands to make a stronger conclusion. When using ephrin-B1, growth cone collapse in FBD1 WT is not significant comparing to Fc treatment. When using ephrin-B2, growth cone collapse in FBD1 WT is not as significant as it is in FBD1 mut group (*** versus **). We interpret this as meaning that the EPHB2-mediated growth cone collapse to both ligands is dampened, when we disrupt the EPHB2-MYCBP2 association. The difference between these two ligands might be due to their different affinities for the receptor or signalling kinetics.
- By far the weakest link in this paper is the worm part. I think it's a pity because strengthening this would affect the significance of the finding. First, the authors mention new genes without introducing their relationship to the signaling pathway tested. Second, the textual logics should be strengthened. Finally and most importantly, when the difference between the phenotypic severity is so strong (vab-1 and rpm-1) then I think it's impossible to say anything from the double mutant.
We appreciate the reviewer noting that they appreciate the value and importance of the C. elegans model. The goals of our C. elegans experiments were twofold:
To evaluate genetic interactions between the VAB-1 Eph receptor and known RPM-1 binding proteins. This was not clearly explained in the original manuscript nor was the published precedent for these types of genetic enhancer experiments provided. We have now rectified this by substantially revising the text of the Results C. elegans section starting on line 431 and by adding several citations.
Our C. elegans genetics confirmed that the VAB-1 Eph receptor is not inhibited/degraded by the RPM-1/MYCBP2 ubiquitin ligase complex. We have now revised the text to draw this point out more clearly.
To further address the reviewer’s concerns, we have added a new schematic (Fig. 8A) to show the relationship between the RPM-1 and the RPM-1 binding proteins (FSN-1/FBXO45 and GLO-4/SERGEF) we are testing. We chose FSN-1 because it is part of the RPM-1 ubiquitin ligase complex and we chose GLO-4 because it functions outside the context of RPM-1 ubiquitin ligase signaling via the GLO-1 Rab GTPase to influence late endosomal/lysosomal biogenesis.
Regarding the reviewer’s concern that different penetrance/frequency of defects between rpm-1 mutants and vab-1 mutants means outcomes with vab-1; rpm-1 double mutants cannot be interpreted. We respectfully disagree. An extensive number of published studies have demonstrated that RPM-1 binding proteins have milder phenotypes than rpm-1 mutants and display genetic enhancer effects as double mutants with one another (PMID:17698012, PMID: 22357847, PMID: 25010424, PMID: 24810406). We now make this point much more clearly. While the frequency of axon termination defects in rpm-1 mutants is high it is not completely saturated as the defect is not 100%. Moreover, a major point of the vab-1; rpm-1 double mutants is that they do not have a significant reduction in phenotypic penetrance/frequency. Thus, our system is fully capable of resolving genetic suppression, which did not occur. We now make this point much more carefully and clearly.
To further address the reviewer’s concern, we have softened language about the VAB-1/Eph receptor functioning in the same pathway as RPM-1 throughout the manuscript. While we think this is still the case, because the frequency of axon termination defects is not fully saturated in rpm-1 mutants and defects could potentially become more severe (i.e. the hook might occur closer to the head of the animal rather than in the midbody). Nonetheless, this is not a critical point and we think it is more important to be clear about the two major goals and objectives of our C. elegans experiments. We hope the reviewer agrees that our rationale, logic and conclusions are more clearly and accurately drawn in the revised paper.
-
eLife assessment
This valuable study identifies an Ephrin type-B Receptor 2 (EPHB2) interactor, MYCBP2, as a potential regulator of EPHB2 stability and function. In contrast to expectations, based on MYCBP2 function in the ubiquitin pathway, loss of function of MYCBP2 resulted in less EPHB2 receptor and defective EPHB2 function. The paper is supported by a largely convincing set of biochemical, cell culture and in vivo experiments.
-
Reviewer #1 (Public Review):
The Eph receptor tyrosine kinase family plays a critical function in multiple physiological and pathophysiological processes. Hence, understating the regulation of these receptors is highly important question. Through extensive experiments in cell lines and cultured neurons Chang et.al show that the signaling hub protein, MYCBP2 positively regulates the overall stability of a specific member of the family, EPHB2, and by that the cellular response to ephrinBs.
Overall, this work sheds light on the divergent in the regulatory mechanisms of the Eph receptors family. Although the physiological importance of this new regularly mechanism in mammals awaits to be discovered, the authors provide genetic evidence using C.elegans that it is evolutionarily conserved. -
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) is also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed …
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) is also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed that FBXO45 within a ternary complex increased the stability of the EphB2/MYCBP2 complex.
To define the interacting domains, they used clearly designed swapping experiments to show that the extracellular and transmembrane domains are necessary and sufficient for the formation of the ternary complex.
Using a cellular contraction assay, the authors showed the necessity of MYCBP2 in mediating the cytoskeletal response of EphB2 forward signalling. Furthermore, they used the technically challenging stripe assay of alternating lanes of ephrinB-Fc and Fc to show that also in this migration-based essay MYCBP2 is required for EphB mediated differential migration pattern.
MYCBP2 in addition is necessary to stabilize EphB2, that is in the absence of MYCBP2, EphB2 is degraded in the lysosomal pathway.
Interestingly, the third protein in this complex, Fbxo45, was further characterized by overexpression of the domain of MYCBP2, known to interact with Fbxo45. Here the authors showed that this approach led to the disruption of the EphB2 / MYCBP2 complex, and also abolished the ephrinB mediated activation of EphB2 receptors and their differential outgrowth on ephrinB2-Fc / Fc stripes.
Finally, the authors demonstrated an in vivo function of this complex using another model system, C elegans where they were able to show a genetic interaction.
Data show in a nice set of experiments a novel level of EphB2 forward signalling where a ternary complex of this receptor with multifunctional MYCBP2 and Fbxo45 controls the activity of EphB2, allowing a further complex regulation of this important receptors. Additionally, the authors challenge pre-existing concepts of the function of MYCBP2 which might open up novel ways to think about this protein.
Of interest is this work also in terms of development of the retinotectal projection in zebrafish where MYCBP2/highwire plays a crucial role, and thus might lead to a better understanding of patterning along the DV axis, for which it is known that EphB family members are crucial.Overall, the experiments are classical experiments of co-immunoprecipitations, swapping experiments, collapse assays, and stripe assays which all are well carried out and are convincing.
-
Reviewer #3 (Public Review):
In this improved version of the manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, suggesting the involvement of additional proteins as MYCBP2 is thought to be a cytoplasmic protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 …
Reviewer #3 (Public Review):
In this improved version of the manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, suggesting the involvement of additional proteins as MYCBP2 is thought to be a cytoplasmic protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 as a ubiquitin E3 ligase, it actually positively regulates EPHB2 protein stability, and function.
The strength of this manuscript is the extensive biochemical analysis of the EPHB2/MYCBP2/FBXO43 interactions. The vast majority of the conclusions supported by the data.
The attempt to extend the study to an in vivo animal using the worm is important, however the additive insight is, unfortunately, minimal.
-
-
eLife assessment
This valuable study provides solid evidence that the stability and function of the Eph-B2 receptor (EPHB2) are affected by interactions with the multifunctional MYCBP2 and Fbxo45 proteins, extending our knowledge of how members of this key family of receptors are regulated, in particular in regard to their forward signaling. The biochemical binding evidence is generally convincing, but the evidence from the C. elegans experiments is still incomplete.
-
Reviewer #1 (Public Review):
The Eph receptor tyrosine kinase family plays a critical function in multiple physiological and pathophysiological processes. Hence, understating the regulation of these receptors is a highly important question. Through extensive experiments in cell lines and cultured neurons, Chang et.al show that the signaling hub protein, MYCBP2 positively regulates the overall stability of a specific member of the family, EPHB2, and by that the cellular response to ephrinBs. Overall, this work sheds light on the divergence in the regulatory mechanisms of the Eph receptors family. The physiological importance of this new regular mechanism awaits discovery.
-
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type-specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed that …
Reviewer #2 (Public Review):
Members of the EphB family of tyrosine kinase receptors are involved in a multitude of diverse cellular functions, ranging from the control of axon growth to angiogenesis and synaptic plasticity. In order to provide these diverse functions, it is expected that these receptors interact in a cell-type-specific manner with a diverse variety of downstream signalling molecules.
The authors have used proteomics approaches to characterise some of these molecules in further detail. This molecule, myc-binding protein 2 (MYCBP2) also known as highwire, has been identified in the context of establishment of neural connectivity. Another molecule coming up on this screen was identified as FBXO45.
The authors use classical methods of co-IP to show a kinase-independent binding of MYCBP2 to EphB2. They further showed that FBXO45 within a ternary complex increased the stability of the EphB2/MYCBP2 complex.
To define the interacting domains, they used clearly designed swapping experiments to show that the extracellular and transmembrane domains are necessary and sufficient for the formation of the ternary complex.
Using a cellular contraction assay, the authors showed the necessity of MYCBP2 in mediating the cytoskeletal response of EphB2 forward signalling. Furthermore, they used the technically challenging stripe assay of alternating lanes of ephrinB-Fc and Fc to show that also in this migration-based essay MYCBP2 is required for EphB mediated differential migration pattern.
MYCBP2 in addition is necessary to stabilize EphB2, that is in the absence of MYCBP2, EphB2 is degraded in the lysosomal pathway.
Interestingly, the third protein in this complex, Fbxo45, was further characterized by overexpression of the domain of MYCBP2, known to interact with Fbxo45. Here the authors showed that this approach led to the disruption of the EphB2 / MYCBP2 complex, and also abolished the ephrinB-mediated activation of EphB2 receptors and their differential outgrowth on ephrinB2-Fc / Fc stripes.
Finally, the authors demonstrated an in vivo function of this complex using another model system, C elegans where they were able to show a genetic interaction.
Data shows in a nice set of experiments a novel level of EphB2 forward signalling where a ternary complex of this receptor with multifunctional MYCBP2 and Fbxo45 controls the activity of EphB2, allowing a further complex regulation of this important receptor. Additionally, the authors challenge pre-existing concepts of the function of MYCBP2 which might open up novel ways to think about this protein.
Of interest is this work also in terms of the development of the retinotectal projection in zebrafish where MYCBP2/highwire plays a crucial role, and thus might lead to a better understanding of patterning along the DV axis, for which it is known that EphB family members are crucial.
Overall, the experiments are classical experiments of co-immunoprecipitations, swapping experiments, collapse assays, and stripe assays which all are well carried out and are convincing.
-
Reviewer #3 (Public Review):
In this manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 as a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by the binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, something that is surprising given the fact that MYCBP2 is an intracellular protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 as a ubiquitin E3 ligase, it …
Reviewer #3 (Public Review):
In this manuscript, Chang et al set out to find direct interactions with the Eph-B2 receptor, as our knowledge of its function/regulation is still incomplete. Using proteomic analysis of Hela cells expressing EPHB2, they identified MYCBP2 as a potential binder, which they then confirm using extensive biochemical analyses, an interaction that seems to be negatively affected by the binding of ephrin-B2 (but not B1). Furthermore, they find that FBXO45, a known MYCBP2 interaction, strongly facilitates its binding to EPHB2. Intriguingly, these interactions depend on the extracellular domains of EPHB2, something that is surprising given the fact that MYCBP2 is an intracellular protein. Finally, they find that, in contrast to what could be expected given the known function of MYCBP2 as a ubiquitin E3 ligase, it actually positively regulates EPHB2 protein stability, and function.
The strength of this manuscript is the extensive biochemical analysis of the EPHB2/MYCBP2/FBXO43 interactions. Most of the conclusions are warranted although I do not understand the physiological interpretation of how these proteins could interact in the extracellular space.
The attempt to extend the study to an in vivo animal using the worm is important. However, I find the results in the worm confusing and overly interpreted in their current form.
-
