Substrate evaporation drives collective construction in termites
Curation statements for this article:-
Curated by eLife

eLife assessment
This valuable study investigates the environmental drivers behind termite construction, focusing, in particular, on pellet deposition behavior, with the conclusion that termites likely sense curvature indirectly through substrate evaporation. The findings reconcile discrepancies between previous studies through experimental and computational approaches. While the strength of the evidence supporting these claims is compelling, the authors do not discuss how their results affect our understanding of insect nest construction or animal-built structures more broadly.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
- @lana's saved articles (lana)
Abstract
Termites build complex nests which are an impressive example of self-organization. We know that the coordinated actions involved in the construction of these nests by multiple individuals are primarily mediated by signals and cues embedded in the structure of the nest itself. However, to date there is still no scientific consensus about the nature of the stimuli that guide termite construction, and how they are sensed by termites. In order to address these questions, we studied the early building behavior of Coptotermes gestroi termites in artificial arenas, decorated with topographic cues to stimulate construction. Pellet collections were evenly distributed across the experimental setup, compatible with a collection mechanism that is not affected by local topography, but only by the distribution of termite occupancy (termites pick pellets at the positions where they are). Conversely, pellet depositions were concentrated at locations of high surface curvature and at the boundaries between different types of substrate. The single feature shared by all pellet deposition regions was that they correspond to local maxima in the evaporation flux. We can show analytically and we confirm experimentally that evaporation flux is directly proportional to the local curvature of nest surfaces. Taken together, our results indicate that surface curvature is sufficient to organize termite building activity and that termites likely sense curvature indirectly through substrate evaporation. Our findings reconcile the apparently discordant results of previous studies.
Article activity feed
-

-
-
-

Author Response
The following is the authors’ response to the previous reviews.
This paper now provides a convincing presentation of valuable results of the drivers of nest construction for one termite species, and they briefly discuss possible relevance to other termite species. However, the authors have not yet addressed how their results may be important outside the field of termite nest construction. I could imagine the significance of the paper being elevated to important if there is a broader discussion about the impact of this work, e.g., the relevance of the results, the approach, and/or next steps to related fields outside of termite nest construction.
Reading our manuscript again, we have to agree with the reviewer that we mostly focused the discussion of our results in the context of termite construction, without attempting …
Author Response
The following is the authors’ response to the previous reviews.
This paper now provides a convincing presentation of valuable results of the drivers of nest construction for one termite species, and they briefly discuss possible relevance to other termite species. However, the authors have not yet addressed how their results may be important outside the field of termite nest construction. I could imagine the significance of the paper being elevated to important if there is a broader discussion about the impact of this work, e.g., the relevance of the results, the approach, and/or next steps to related fields outside of termite nest construction.
Reading our manuscript again, we have to agree with the reviewer that we mostly focused the discussion of our results in the context of termite construction, without attempting to generalise to other systems. To some extent we still defend this choice, as we prefer not to make too many claims on the relevance of our results beyond what we can reasonably support with our own experimental results. However, we thought that it would be appropriate – as suggested by the reviewer – to add at least one paragraph to indicate how our results could be extrapolated to other systems. This new paragraph is now at the end of the discussion section.
Here we elaborate a bit further on this point: first of all, while termites certainly build the most complex structures found in the natural world, there aren’t many other animals that are capable of collectively building complex structures. Typically, collective building activity is limited to highly social (typically eusocial) animals, but other social insects, such as ants and wasps, are phylogenetically distant from termites, their nests are often different (the large majority of ant nests only comprise excavated galleries with little construction, while wasp nests tend to comprise multiple repeated patterns that could be produced from stereotyped individual behaviour). Because of these differences, drawing a comparison between the mechanisms that regulate termite architecture and those that regulate other forms of animal architecture would be too speculative. One domain, however, where similar mechanisms to those that we describe here could operate is that of pattern formation at the cellular and tissue level, where surface curvature was shown to drive different phenomena from cell migration to tissue growth. A comment on this is now added in the manuscript at the very end of the discussion.
Similarly, on a related note, as someone not directly in the field of termite nest construction but wanting to understand the system (and the results) presented here in a broader context, I found the additional information about species and natural habitat very helpful and interesting, though I was rather disappointed to find it relegated to supplementary material where most readers will not see it.
We considered this suggestion to present more information about the natural nesting habits of the termites that we study into the main text, but eventually we decided to leave it as supplementary only. We feel that the nesting habits of the termites that we studied here are not too central to the problem that we want to focus on, of how they coordinate their building activity. In fact, there is a large variety of nesting habits across termite genera and species, but we believe that, at a basic level, the mechanisms that we describe here would also apply to species with different nesting habits, because our results are consistent with what is described in the scientific literature for other termite species. As our introduction is already a bit long, we left this description of Coptotermes nesting habits in the supplementary material, where, hopefully, it will still be accessible and useful to readers interested in finding this information.
When providing responses to reviewers, please directly address the reviewers’ comments point-by-point rather than summarizing comments and responding to summaries.
We apologize for our previous way to respond to comments and thanks the reviewer for his remark as we learn to navigate through the eLife reviewing system (where some comments are repeated in the overall assessment and in the feed-back of individual reviewers).
Figure 2 colors: Panels A and E and maybe B do not seem colorblind-friendly. I suggest modifying the colormaps to address this.
We have changed the colormaps of figures A,B and E which are now colorblind-friendly.
Line 180: This system is not in equilibrium. Perhaps the authors mean "steady-state?" I suggest reviewing language to ensure that the correct technical terms are used.
We have now corrected this.
-
eLife assessment
This valuable study investigates the environmental drivers behind termite construction, focusing, in particular, on pellet deposition behavior, with the conclusion that termites likely sense curvature indirectly through substrate evaporation. The findings reconcile discrepancies between previous studies through experimental and computational approaches. While the strength of the evidence supporting these claims is compelling, the authors do not discuss how their results affect our understanding of insect nest construction or animal-built structures more broadly.
-
Joint Public Review:
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
Through simulations and ingenious experiments the authors have shown how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously developed model for termite construction, to highlight and verify their findings. On my …
Joint Public Review:
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
Through simulations and ingenious experiments the authors have shown how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously developed model for termite construction, to highlight and verify their findings. On my first pass of the manuscript I felt the authors were missing an experiment: an array of humidity probes to measure evaporation in the three spatial dimensions and over time. Technologically such an experiment is not out of reach, but the author's alternative (a substrate made with a saline solution and later measuring the salt deposits on the surface) was a very ingenious low tech solution to the problem.
The authors agree that future experiments should tackle finely controlled humidity levels and curvature in order to have a more quantitative measure termite behaviour, but the work done so far is more than sufficient to justify their current claims.
In the revised text, the authors have added more clarity into different biological systems in which these results could be applied. Perhaps what it would have been beneficial to also add more information on how the resulting algorithms of constructions can be used in swarm robotics with collective construction, both macro and micro, but I acknowledge that the style of the paper does focus more on the biological aspects
The results presented here are so far the best attempt on characterizing multiple cues that induce termite construction activity, and that possibly unifies the different hypothesis presented in the last 8 years into a single factor, resulting into a valuable addition to the field. More importantly, even if these results come from different species of termites than some of the previous works, they are relatable and seem to be mostly consistent, improving the strength of the author's claims.
-
-
Author Response
The following is the authors’ response to the original reviews.
We would like to thank you and the two reviewers for their constructive feed-back on our manuscript entitled: "Substrate evaporation drives collective construction in termites".
Here, we submit a revised version in which -we believe- we fill the missing details identified by the reviewers and we clarify the presentation of our results.
From the eLife assessment we can identify a few main points that the reviewers found unclear or not well developed in our previous manuscript:
• Insufficient details about computer simulation models. Is the match between simulations and experiments qualitative or quantitative?
• Request for clarifications related to the wall stimulus: is evaporation stronger at the high-curvature wall corners or similar along all the wall …
Author Response
The following is the authors’ response to the original reviews.
We would like to thank you and the two reviewers for their constructive feed-back on our manuscript entitled: "Substrate evaporation drives collective construction in termites".
Here, we submit a revised version in which -we believe- we fill the missing details identified by the reviewers and we clarify the presentation of our results.
From the eLife assessment we can identify a few main points that the reviewers found unclear or not well developed in our previous manuscript:
• Insufficient details about computer simulation models. Is the match between simulations and experiments qualitative or quantitative?
• Request for clarifications related to the wall stimulus: is evaporation stronger at the high-curvature wall corners or similar along all the wall edge? Why is there less consistency in the experimental results with the wall stimulus, with a minority of wall experiments in which something different happens?
• Quantitative estimation of the humidity gradients in our experimental setup.
• "Confirmation" that termites can sense humidity gradients of magnitude and scale comparable with those encountered in our experiments.
• Request for additional background information about the considered termite species and their construction habits.
The reviewers also made a number of interesting suggestions and other comments:
• Suggestion of possible explanations and interpretations for a purported discrepancy with a previous work by Calovi and collaborators.
• Suggestion of alternative experimental approaches (array of probes, alternative experimental setups).
We address all these points below.
Details about computer simulation models
There are two different types of computer simulations in our experiments: 1. simulations of evaporation on the initial structure, and 2. simulations of structure growth based on curvature.
- Simulations of evaporation We recall that these simulations rely on the hypothesis that humidity transport happens in a diffusive way, that is evaporation rate is proportional to the humidity gradient. New details on the implementation of these diffusive simulations are now added in section S.VI. We also adapted figures 4A and 4B which are now expressed in units more comparable to the expected humidity field in experiments. Essentially, we show that the model under-estimates the absolute magnitude of the humidity gradient |∇ℎ| in our setup while it correctly predicts the relative importance of the same field across the topography.
First, it is instructive to report the value of |∇ℎ| predicted by diffusive simulations with the bottom boundary at 100% humidity (like the clay disk), and the top boundary of the simulation box at 70% like our experimental room. Note that, at a given temperature, relative humidity and absolute humidity are proportional, so we will assume here that temperature is constant and always refer to relative humidity. Thus, humidity gradient will be measured in 𝑚𝑚−1 exactly like curvature. One than has:
• flat disk, |∇ℎ| ∼0.01mm−1
• wall tips, |∇ℎ| ∼0.13mm−1
• wall top edge |∇ℎ| ∼0.1mm−1
• pillar tips |∇ℎ| ∼0.19mm−1,
First we remark, that the value of |∇ℎ| on the flat portion of the disk is 10 times smaller of the estimation |∇ℎ|0 ∼0.5mm−1 of the same quantity in our experiments, which is now given in the manuscript and discussed in a specific paragraph below. This discrepancy is due to the fact that our simulations overestimate the size of the diffusive region (i.e. the simulation box) to 18mm while we expect the diffusive layer to be much thinner (i.e. 𝛿 ∼2mm). Note also that, as in all diffusive problems, the humidity gradient on any point of the bottom boundary (i.e. on the clay surface) depends on the distance of that point from the top boundary, for example the closer are the boundaries the stronger is the gradient. This is a very general feature of diffusive problems: the gradient of the diffusing field depends on the distance from the boundaries, where the value of the field is given. Note also that, in principle, the size of the simulation box does not only affect the overall magnitude of the humidity gradient but also its shape. However, one observes that in our simulations the topographic cues are only 30% closer to the top boundary compared to the flat, bottom, surface, but the local gradient is 10 to 20 times larger. This evidence suggests that the ’curvature’ effect is much stronger than the ’distance’ effect, and supports the fact that our approximation does not affect in a significant way the estimation of the relative importance of the humidity gradient at the bottom surface. We then conclude that our diffusive simulations do not provide a correct estimation of the order of magnitude of |∇ℎ|, but well capture its relative variations across the topography.
- Structure growth based on curvature. As observed by the reviewer, the dynamical simulations included here refer to a model that was developed in a previous study, thus we chose to not include all the details of the simulations in the present one. At this stage, that model is still phenomenological: for example we cannot provide a physical estimation of the dimensionless parameter 𝑑 which controls the typical size of the structure produced by the simulations of the model. Thus in principle, the comparisons with real experiments cannot be other than "qualitative". Indeed, to push such a comparison further is not necessarily of interest, given the minimal and mean field character of our model, and the extreme complexity of the natural system which is studied here. However, our experimental setup was specifically designed to overcome this limit, which is designing topographies where the curvature cues where modulated in a way which is almost discrete, with flat regions, and regions where curvature is strong ’for termites’, i.e. the curvature radius is of the order of termite body size. Our experimental results greatly validate our choice because deposition patterns also show an almost ’discrete’ shape, with specific regions attracting most of the depositing actions. Thus, we claim that the significance of the agreement is strong, and we suggest that when stimuli and response both behave in a quasi-discrete manner, the difference between qualitative and quantitative is not well defined. Finally, we recall that in all the discussion above curvature and humidity gradient can be exchanged, as we already pointed out in the manuscript. Consistently, the humidity gradient show a strong variation between the curved regions and the flat ones.
Results with the wall stimulus One important point coming out from the reviews is that we did not clearly present the results with the wall stimulus. These concerns are best summarized by a comment from reviewer 2, who states: “evaporation rates seem inconclusive in the wall geometry, yet the termites still deposit material at the high-curvature wall corners”.
We acknowledge that the interpretation of results of experiments with the wall stimulus must address three key points: 1- Salt deposition experiment are inconclusive in showing variation of the evaporation rate, across the top of the wall; 2- A portion (4/11) of termite experiments do not show a clear pellet deposition pattern by termites; 3- Conversely, in the remaining portion (7/11), most experiments still show a clear pellet deposition on the corners of the wall, in spite of small differences in evaporation between the corners and the top edge (like in our Fig. 3B). These points are now addressed in the manuscript and discussed below.
The variation of the humidity gradient between the corners of the wall, and the wall’s top edge is relatively small while both are regions of relatively high curvature and higher evaporation as compared to the the flat surface of the clay disk. We now report precise values of the humidity gradient from numerical simulations, as discussed above. These indicate that humidity gradient at the wall corners and upper edge is respectively 10 and 7 times larger than on the flat bottom, but evaporation at the wall tips is only 0.3 times larger than on the wall upper edge.
Experiments with the saline solution qualitatively confirm the same result of an evaporation pattern more evenly distributed on the wall stimulus (point 1) than on the pillars.
Taken together, these results might explain why not all wall experiments end up with depositions at the tips (point 2): simply, in the wall experiments the relative importance of the deposition cue between tips and wall upper edge is not high enough to always guide termite behavior in a deterministic way.
But we should also point to the fact that the evaporation simulations presented in figure 4 and the experiments with the saline solution both reflect the humidity field on the clay templates before termite construction has started. As soon as termites start adding pellets to the wall, effectively starting to build a pillar, the humidity gradient will be reinforced at the locations of pellet deposition, and a self-reinforcing process is initiated, similar to our dynamical simulations based on local curvature. This explains why eventually termite activity can result in clear and localized depositions (point 3) also with the wall stimulus.
Incidentally, we would like to include here another consideration: the nest of Coptotermes termites comprise a “scaffold” with multiple interconnected pillars. In other termite genera, the prevalent nest structure is one made by surfaces, rather than pillars, such as in Nasutitermes nests, Apicotermes, Psammotermes, or again some fungus growing structures in Macrotermes and Synacanthotermes). The fact that the wall stimulus presents some potential to stimulate construction everywhere on its edge is intriguing as it might provide some cues on the construction of different nest architectures.
Quantitative estimation of the humidity gradient in our setup The moisture gradients in our experiments and simulations was only presented in a non-quantitative manner, because we were mainly interested in identifying locations of high and low evaporation. But, combining scaling arguments already discussed in S.IX and the the results of our evaporation simulations, one can produce a lower boundary for the magnitude of the humidity gradient |∇ℎ|, predict its higher value at key positions on our setup, and compare it with humidity variations experienced by termites in their natural environment. These considerations are now included in the manuscript and discussed below.
First, we define a reference value |∇ℎ|0 for the humidity gradient on the (flat) clay disk, which can be estimated using the boundary layer thickness 𝛿 ∼2mm (see section IX.A of the SI) and the variation of relative humidity Δℎ between the clay disk surface and the exterior which was Δℎ =30% (the difference between the fully wetted substrate, and room air humidity at 70% saturation). Note that |∇ℎ|0 constitutes a lower boundary for the expected values of the humidity gradient in our setup, as confirmed by our experiments with saline solution. We can then write:
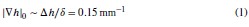
Next, the results of diffusive simulations shown in figure 4A and 4B indicate that the humidity gradient at highly curved regions of the topographic cues is at least 10 times larger than |∇ℎ|0 which allows to estimate an upper boundary for |∇ℎ| in our experimental setup, say |∇ℎ|𝑚𝑎𝑥 ∼1mm−1. Humidity sensing capabilities of termites Our hypothesis that humidity gradients could guide termite building behavior implicitly assumes that termites can sense humidity gradients comparable with those existing in our experiments.
Humidity is important to all termites because of their small size and unsclerotized body. Coptotermes termites in particular are wetwood termites that can only survive in high-humidity environments such as moist wood or soil. It is well documented that coptotermes termites (like other termites and cockroaches) have humidity receptors in their antennae, and behavioral studies indicate that they can discriminate between chambers with different humidity content.
For example, a study by Gautam and Henderson (2011, Environmental entomology, 40:1232) provided chambers with different relative humidity and, after 12 hours, almost all termites were in the highest humidity chamber (98% RH), leaving the other chambers with 75% or less RH empty. These results (which are similar also to other results testing termite response to chambers with different soil moisture) indicate that -given a sufficient amount of time- termites can detect a difference of humidity from 75% to 98% over a spatial scale of centimeters.
The quantitative estimation of the humidity gradient described above indicates that in our experimental setup termites can experience humidity variations of 15% over a distance of only 1mm and even shorter, while the length of a single termite antenna is about 1.5 mm.
In other words, the humidity gradients that we estimate for our experiments are well above those that termites were able to discriminate in previous experiments. Future experiments should aim to test the exact limits of resolution of the humidity-sensing ability of termites (e.g. in an environment where humidity is close to 100% everywhere), and the mechanisms how they sense the gradient (e.g. comparing information from the two antennae, or by integrating humidity information over time).
By definition, |∇ℎ|0 corresponds to a variation of humidity between a fully saturated atmosphere (i.e. 100%), comparable to the nest interior, and a "humid" atmosphere (i.e. 70%) comparable to the natural environment where termites live (say the nest exterior), occurring over a distance (2mm) which is comparable with their body size.
We can then conclude that even the lower boundary |∇ℎ|0 of the humidity gradient corresponds to an atmosphere variation to which termites must be used, i.e. nest interior vs nest exterior, happening across one body length. If we add that the upper boundary |∇ℎ|𝑚𝑎𝑥 is one order of magnitude higher, it appears extremely unlikely that they could not detect these gradients.
Additional background information about our considered termite species and their construction habits
We have now added some details about the life history and nesting habits of termites in the Coptotermes genus in a new paragraph in section SI. Essentially, these are wetwood termites that nest in moist wood or soil, and their nests present a typical structure comprising a scaffold of interconnected pillars (we now show a picture of a typical structure from one of our lab-reared colonies).
After the initial submission of our manuscript we have also obtained a more precise taxonomic identification of the termites we used, which indicated that our termites are better identified as Coptotermes gestroi than Coptotermes formosanus. The two species are extremely close and can also interbreed in the areas where they co-occur, but in this case C. gestroi is a better match. Hence, we have amended the name in the manuscript and in the supplementary material.
Differences with previous results by Calovi and collaborators
We believe that there is no real discrepancy between our results and those described by Calovi et al. (2019, Phil. Trans. Roy. Soc. B 374:20180374). What they measure-termite aggregation and activity- is similar to what we also observe in our experiments: termites aggregate in concave regions, such as at the base of the wall in our experiments, and they collect pellets at the locations that they visit more often. And, above all, we observe that concavities promote digging activity, which in turns promote aggregation as already observed in previous studies like Green et al. (2017, Proc. Roy. Soc. B 284:20162730). The main difference is that in our analyses we treat separately the three measurements of termite occupancy, pellet collection and pellet deposition, and in this way we identify a role of convexity for pellet deposition.
It is possible that, apart from the differences in language and interpretations between our study and the study by Calovi, there were also real differences in termite building behavior between the two studies that we couldn’t fully appreciate from our own reading of the article by Calovi, but which the reviewer has spotted. The reviewer makes a very interesting suggestion that some of these differences might be due to the different humidity level used in our experiment, compared to the experiment by Calovi and collaborators. Room humidity was high, at around 70% in our experiments. The humidity in Calovi’s experiments was possibly even higher as they performed their experiments in a closed box, but we could not find precise reported information on the humidity level in their publication.
Given that it is not clear that the building behavior in our experiments was qualitatively different from the building behavior in Calovi and collaborators’ experiments, and given that we don’t know the precise humidity value used in Calovi’s experiments (plus, we worked on different termite species that could have different sensitivity to humidity) we decided that -based on the information that we have- we could not meaningfully expand our discussion of similarities and differences with Calovi’s study in our manuscript.
It is clear, though, and we completely agree with the referee on this point, that in light of Calovi’s and our own new results, it would now be extremely interesting if future experiments could characterize termite construction activity across a range of finely controlled air humidity values. Anecdotally, in preliminary experiments we did include some trials in which termites were hosted in a completely closed box, and we observed much reduced construction activity in those conditions. However, the fact that we could not easily track termite activity and pellet collections / depositions in those conditions (because of the box), together with the fact that the building activity itself was reduced, made us to converge towards the open arena experiments that we describe here.
Suggestion of alternative experimental approaches One reviewer made interesting suggestions for alternative experiments, including using an array of humidity probes for measuring humidity, or a different experimental setup -analogous to those used in previous experiments by Bardunias and collaborators-. It is often the case that only at the end of a series of experiments we identify an alternative, and possibly better, way of doing the same experiment. In future, if we have the opportunity to run other similar experiments again, we will likely experiment with these suggestions. When we first designed our own experiments, one of our priorities was to be able to film all termites in the arena at all time, so that potentially we could also study individual termite behavior and task specialization. This partly constrained the type of experimental setups that we could use.
One aspect that clearly emerged from our work and from the revision process is that any future experiments related to this topic should achieve a very precise control of air humidity, and test a wider range of stimuli of more varied and controlled size, humidity and curvature. Since our own experiments were conducted, three of us have moved to different institutions, which imposes practical constraints for us on working on the same termites in a similar way, but the suggestions from the reviewers will be helpful as we are planning our future research.
We hope that the explanations above and the details that we have changed in the manuscript itself have contributed to clarify unclear aspects of our study.
-
eLife assessment
This valuable study investigates the environmental drivers behind termite construction, focusing, in particular, on pellet deposition behavior, with the conclusion that termites likely sense curvature indirectly through substrate evaporation. The findings reconcile discrepancies between previous studies through experimental and computational approaches. Although the strength of the evidence supporting these claims is compelling, the authors do not discuss how their results affect our understanding of insect nest construction or animal-built structures more broadly.
-
Reviewer #1 (Public Review):
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
Through simulations and ingenious experiments the authors have shown how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously developed model for termite construction, to highlight and verify their findings. …
Reviewer #1 (Public Review):
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
Through simulations and ingenious experiments the authors have shown how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously developed model for termite construction, to highlight and verify their findings. On my first pass of the manuscript I felt the authors were missing an experiment: an array of humidity probes to measure evaporation in the three spatial dimensions and over time. Technologically such an experiment is not out of reach, but the author's alternative (a substrate made with a saline solution and later measuring the salt deposits on the surface) was a very ingenious low tech solution to the problem.
The authors agree that future experiments should tackle finely controlled humidity levels and curvature in order to have a more quantitative measure termite behaviour, but the work done so far is more than sufficient to justify their current claims.
The results presented here are so far the best attempt on characterizing multiple cues that induce termite construction activity, and that possibly unifies the different hypothesis presented in the last 8 years into a single factor. More importantly, even if these results come from different species of termites than some of the previous works, they are relatable and seem to be mostly consistent, improving the strength of the author's claims.
-
Reviewer #2 (Public Review):
The revised paper addresses most of my major comments and concerns. The authors have added more detail explaining their model, they have added more background information, citations, and discussion for termite humidity sensing capabilities. With these modifications, this paper now provides a convincing presentation of valuable results of the drivers of nest construction for one termite species, and they briefly discuss possible relevance to other termite species. However, the authors have not yet addressed how their results may be important outside the field of termite nest construction. I could imagine the significance of the paper being elevated to important if there is a broader discussion about the impact of this work, e.g., the relevance of the results, the approach, and/or next steps to related fields …
Reviewer #2 (Public Review):
The revised paper addresses most of my major comments and concerns. The authors have added more detail explaining their model, they have added more background information, citations, and discussion for termite humidity sensing capabilities. With these modifications, this paper now provides a convincing presentation of valuable results of the drivers of nest construction for one termite species, and they briefly discuss possible relevance to other termite species. However, the authors have not yet addressed how their results may be important outside the field of termite nest construction. I could imagine the significance of the paper being elevated to important if there is a broader discussion about the impact of this work, e.g., the relevance of the results, the approach, and/or next steps to related fields outside of termite nest construction. Similarly, on a related note, as someone not directly in the field of termite nest construction but wanting to understand the system (and the results) presented here in a broader context, I found the additional information about species and natural habitat very helpful and interesting, though I was rather disappointed to find it relegated to supplementary material where most readers will not see it.
-
-
eLife assessment
This valuable study investigates the environmental drivers behind termite construction, focusing, in particular, on pellet deposition behavior, with the conclusion that termites likely sense curvature indirectly through substrate evaporation. The findings reconcile discrepancies between previous studies through experimental and computational approaches. The strength of the evidence supporting the claims is solid, although many computational details are still missing, and the are concerns about the repeatability of the results across replicates, the dynamics of evaporation in their experiment, and the termites' ability to sense humidity.
-
Reviewer #1 (Public Review):
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, with independent (and sometimes conflicting) hypotheses, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
The authors managed through simulations and ingenious experiments to show how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can most of the time be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously …
Reviewer #1 (Public Review):
In this manuscript the authors performed experiments and simulations which showed that substrate evaporation is the main driver of early construction in termites. Additionally, these experiments and simulations were designed taking into account several different works, with independent (and sometimes conflicting) hypotheses, so that the current results shine a light on how substrate evaporation is a sufficient descriptor of most of the results seen previously.
The authors managed through simulations and ingenious experiments to show how curvature is extremely correlated with evaporation, and therefore, how results coming from these 2 environmental factors can most of the time be explained through evaporation alone. The authors have continued to use their expertise of numerical simulations and a previously developed model for termite construction, to highlight and verify their findings. On my first pass of the manuscript I felt the authors were missing an experiment: an array of humidity probes to measure evaporation in the three spatial dimensions and over time. Technologically such an experiment is not out of reach, but the author's alternative (a substrate made with a saline solution and later measuring the salt deposits on the surface) was a very ingenious low tech solution to the problem.
One possible missing experiment (and possibly the explanation of the only inconsistency of their results to previous literature) is to perform similar topographical experiments in high humidity chambers, where no humidity, or very low humidity gradients are present. Previous experiments done by Calovi and collaborators in 2019 showed that termite construction activity (without distinguishing digging from deposition) was focused on high curvature (concave) regions, where here the authors have seen higher depositions on convex structures. Despite the difference of "activity" by Calovi 2019 (clearly acknowledged by the authors), another main difference is that the experiments of the 2019 manuscript were performed in a closed chamber with very high humidity, and smooth transitions between regions of positive and negative curvature. Therefore, it stands to reason that the only missing component of the current article, would have been to perform similar experiments with curvature (positive and negative) but under an environment where gradients are reduced to a minimum.
The results presented here are so far the best attempt on characterizing multiple cues that induce termite construction activity, and that also possibly unifies the different hypothesis presented in the last 8 years into a single factor. More importantly, even if these results come from different species of termites than some of the previous works, they are relatable and seem to be mostly consistent, improving the strength of the author's claims.
-
Reviewer #2 (Public Review):
This study investigates the drivers behind termite construction, with a particular focus on the environmental factors that drive pellet deposition. The authors performed experiments and computations in an attempt to disentangle the role of surface curvature, feature elevation, substrate evaporation, and a possible "cement" pheromone on the deposition of soil pellets.
In three different types of experiments, the authors present termites with pre-made, unmarked (pheromone-free) pellets, and they vary pre-existing topographic building cues: some experiments have two pillars, others have a wall, and a third type had no cues. In experiments with topographic cues, the authors find that deposition seems to occur preferentially at the locations of highest curvature (i.e., peaks of pillars and corners of the walls). …
Reviewer #2 (Public Review):
This study investigates the drivers behind termite construction, with a particular focus on the environmental factors that drive pellet deposition. The authors performed experiments and computations in an attempt to disentangle the role of surface curvature, feature elevation, substrate evaporation, and a possible "cement" pheromone on the deposition of soil pellets.
In three different types of experiments, the authors present termites with pre-made, unmarked (pheromone-free) pellets, and they vary pre-existing topographic building cues: some experiments have two pillars, others have a wall, and a third type had no cues. In experiments with topographic cues, the authors find that deposition seems to occur preferentially at the locations of highest curvature (i.e., peaks of pillars and corners of the walls). Complementary experiments and simulations show that locations of highest curvature correspond to locations with highest evaporation rates, at least for pillars. Evaporation rates seem inconclusive in the wall geometry, yet the termites still deposit material at the high-curvature wall corners. The authors conclude that: (1) no "cement" pheromone is needed for construction, (2) that depositions preferentially occur at locations of high curvature (all experiments for pillars, 7 out of 11 experiments for walls), and (3) that evaporation (which is fastest at places of highest curvature, at least for pillars) drives deposition. The experiments and results seem sound and interesting, but some of the interpretations need more justification. For instance, why conclude that evaporation drives construction when there is not a measurable difference in evaporation rate across the wall geometry?
The authors also perform simulations (developed in a previous publication) that agree with their experimental observation that deposition occurs preferentially at locations of high curvature. However, there is not enough detail provided about the simulation to understand the degree to which simulation and experiment agree (e.g., is the agreement qualitative or quantitative?) as well as the significance of the agreement. The authors should provide additional details about the setup and mechanics of the simulation, the outputs and how they connect to experiments, and potential limitations of results/connections to the experimental system. Finally, more background about this termite species would be helpful in putting these results into context. For instance, what is known about the natural habitat and conditions, and natural nest locations and structures? What are (or might be, depending on what is known) the potential abilities/benefits for these animals to sense humidity gradients, and why building at these locations could benefit the animals?
-
