β-cell intrinsic dynamics rather than gap junction structure dictates subpopulations in the islet functional network
Curation statements for this article:-
Curated by eLife

eLife assessment
The manuscript describes a set of detailed modeling and experimental studies to disentangle the respective roles of gap junctional electrical vs. metabolic coupling mechanisms in the synchronization of islet activity. This is of interest due to the importance of islet synchronization and generally islet network properties in the regulation of insulin secretion from the pancreas. The significance of the findings was judged to be mostly important and the strength of evidence provided was judged to be mostly solid overall.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Diabetes is caused by the inability of electrically coupled, functionally heterogeneous β-cells within the pancreatic islet to provide adequate insulin secretion. Functional networks have been used to represent synchronized oscillatory [Ca 2+ ] dynamics and to study β-cell subpopulations, which play an important role in driving islet function. The mechanism by which highly synchronized β-cell subpopulations drive islet function is unclear. We used experimental and computational techniques to investigate the relationship between functional networks, structural (gap junction) networks, and intrinsic β-cell dynamics in slow and fast oscillating islets. Highly synchronized subpopulations in the functional network were differentiated by intrinsic dynamics, including metabolic activity and K ATP channel conductance, more than structural coupling. Consistent with this, intrinsic dynamics were more predictive of high synchronization in the islet functional network as compared to high levels of structural coupling. Finally, dysfunction of gap junctions, which can occur in diabetes, caused decreases in the efficiency and clustering of the functional network. These results indicate that intrinsic dynamics rather than structure drive connections in the functional network and highly synchronized subpopulations, but gap junctions are still essential for overall network efficiency. These findings deepen our interpretation of functional networks and the formation of functional subpopulations in dynamic tissues such as the islet.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
Briggs et al use a combination of mathematical modelling and experimental validation to tease apart the contributions of metabolic and electronic coupling to the pancreatic beta cell functional network. A number of recent studies have shown the existence of functional beta cell subpopulations, some of which are difficult to fully reconcile with established electrophysiological theory. More generally, the contribution of beta cell heterogeneity (metabolism, differentiation, proliferation, activity) to islet function cannot be explained by existing combined metabolic/electrical oscillator models. The present studies are thus timely in modelling the islet electrical (structural) and functional networks. Importantly, the authors show that metabolic coupling primarily drives the islet …
Author Response
Reviewer #1 (Public Review):
Briggs et al use a combination of mathematical modelling and experimental validation to tease apart the contributions of metabolic and electronic coupling to the pancreatic beta cell functional network. A number of recent studies have shown the existence of functional beta cell subpopulations, some of which are difficult to fully reconcile with established electrophysiological theory. More generally, the contribution of beta cell heterogeneity (metabolism, differentiation, proliferation, activity) to islet function cannot be explained by existing combined metabolic/electrical oscillator models. The present studies are thus timely in modelling the islet electrical (structural) and functional networks. Importantly, the authors show that metabolic coupling primarily drives the islet functional network, giving rise to beta cell subpopulations. The studies, however, do not diminish the critical role of electrical coupling in dictating glucose responsiveness, network extent as well as longer-range synchronization. As such, the studies show that islet structural and functional networks both act to drive islet activity, and that conclusions on the islet structural network should not be made using measures of the functional network (and vice versa).
Strengths:
- State-of-the-art multi-parameter modelling encompassing electrical and metabolic components.
- Experimental validation using advanced FRAP imaging techniques, as well as Ca2+ data from relevant gap junction KO animals.
- Well-balanced arguments that frame metabolic and electrical coupling as essential contributors to islet function.
- Likely to change how the field models functional connectivity and beta cell heterogeneity.
Weaknesses:
- Limitations of FRAP and electrophysiological gap junction measures not considered.
- Limitations of Cx36 (gap junction) KO animals not considered.
- Accuracy of citations should be improved in a few cases.
We thank reviewer 1 for their positive comments, including the many strengths in the approaches, arguments and impact. We do note the weaknesses raised by the reviewer and have addressed them following the comments below.
We would like to also note that when we refer to metabolic activity driving the functional network, we are not referring to metabolic coupling between beta cells. Rather we mean that two cells that show either high levels of metabolic activity (glycolytic flux) or that show similar levels metabolic activity will show increased synchronization and thus a functional network edge as compares to cells with elevated gap junction conductance. Increased metabolic activity would likely generate increased depolarizing currents that will provide an increased coupling current to drive synchronization; whereas similar metabolic activity would mean a given coupling current could more readily drive synchronized activity. We have substantially rewritten the manuscript to clarify this point.
Reviewer #2 (Public Review):
In their present work, Briggs et al. combine biophysical simulations and experimental recordings of beta cell activity with analyses of functional network parameters to determine the role played by gap-junctional coupling, metabolism, and KATP conductance in defining the functional roles that the cells play in the functional networks, assess the structure-function relationship, and to resolve an important current open question in the field on the role of so-called hub cells in islets of Langerhans.
Combining differential equation-based simulations on 1000 coupled cells with demanding calcium, NAPDH, and FRAP imaging, as well as with advanced network analyses, and then comparing the network metrics with simulated and experimentally determined properties is an achievement in its own right and a major methodological strength. The findings have the potential to help resolve the issue of the importance of hub cells in beta cell networks, and the methodological pipeline and data may prove invaluable for other researchers in the community.
However, methodologically functional networks may be based on different types of calcium oscillations present in beta cells, i.e., fast oscillations produced by bursts of electrical activity, slow oscillations produced by metabolic/glycolytic oscillations, or a mixture of both. At present, the authors base the network analyses on fast oscillations only in the case of simulated traces and on a mixture of fast and slow oscillations in the case of experimental traces. Since different networks may depend on the studied beta cell properties to a different extent (e.g., fast oscillation-based networks may, more importantly, depend on electrical properties and slow oscillationbased networks may more strongly depend on metabolic properties), it is important that in drawing the conclusions the authors separately address the influence of a cell's electrical and metabolic properties on its functional role in the network based on fast oscillations, slow oscillations, or a mixture of both.
We thank reviewer 2 for their positive comments, including addressing the importance of this study as it pertains to islet biology and acknowledging methodological complexities of this study. We also thank the reviewer for their careful reading and providing useful comments. We have integrated each comment into the manuscript. Most importantly, we have now extended our analysis to both fast and slow oscillations by incorporating an additional mathematical model of coupled slow oscillations and performing additional experimental analysis of fast, slow, and mixed oscillations.
Reviewer #3 (Public Review):
Over the past decade, novel approaches to understanding beta cell connectivity and how that contributes to the overall function of the pancreatic islet have emerged. The application of network theory to beta cell connectivity has been an extremely useful tool to understand functional hierarchies amongst beta cells within an islet. This helps to provide functional relevance to observations from structural and gene expression data that beta cells are not all identical.
There are a number of "controversies" in this field that have arisen from the mathematical and subsequent experimental identification of beta "hub" cells. These are small populations of beta cells that are very highly connected to other beta cells, as assessed by applying correlation statistics to individual beta cell calcium traces across the islet.
In this paper Briggs et al set out to answer the following areas of debate:
They use computational datasets, based on established models of beta cells acting in concert (electrically coupled) within an islet-like structure, to show that it is similarities in metabolic parameters rather than "structural" connections (ie proximity which subserves gap junction coupling) that drives functional network behaviour. Whilst the computational models are quite relevant, the fact that the parameters (eg connectivity coefficients) are quite different to what is measured experimentally, confirm the limitations of this model. Therefore it was important for the authors to back up this finding by performing both calcium and metabolic imaging of islet beta cells. These experimental data are reported to confirm that metabolic coupling was more strongly related to functional connectivity than gap junction coupling. However, a limitation here is that the metabolic imaging data confirmed a strong link between disconnected beta cells and low metabolic coupling but did not robustly show the opposite. Similarly, I was not convinced that the FRAP studies, which indirectly measured GJ ("structural") connections were powered well enough to be related to measures of beta cell connectivity.
The group goes on to provide further analytical and experimental data with a model of increasing loss of GJ connectivity (by calcium imaging islets from WT, heterozygous (50% GJ loss), and homozygous (100% loss). Given the former conclusion that it was metabolic not GJ connectivity that drives small world network behaviour, it was surprising to see such a great effect on the loss of hubs in the homs. That said, the analytical approaches in this model did help the authors confirm that the loss of gap junctions does not alter the preferential existence of beta cell connectivity and confirms the important contribution of metabolic "coupling". One perhaps can therefore conclude that there are two types of network behaviour in an islet (maybe more) and the field should move towards an understanding of overlapping network communities as has been done in brain networks.
Overall this is an extremely well-written paper which was a pleasure to read. This group has neatly and expertly provided both computational and experimental data to support the notion that it is metabolic but not "structural" ie GJ coupling that drives our observations of hubs and functional connectivity. However, there is still much work to do to understand whether this metabolic coupling is just a random epiphenomenon or somehow fated, the extent to which other elements of "structural" coupling - ie the presence of other endocrine cell types, the spatial distribution of paracrine hormone receptors, blood vessels and nerve terminals are also important.
We thank reviewer 3 for their positive comments, including the methodology, writing style, and the importance of this paper to the broader islet community. We thank the reviewer for their very in-depth and helpful comments. We have addressed each comment below and made significant changes to the manuscript according. We conducted more FRAP experiments and separated results into slow, fast, and mixed oscillations. We included analysis of an additional computational model that simulates slow calcium oscillations. Additionally, we substantially rewrote the paper to clarify that we are not referring to metabolic coupling and speak on the broader implications of network theory and our findings.
Reviewer #4 (Public Review):
This manuscript describes a complex, highly ambitious set of modeling and experimental studies that appear designed to compare the structural and functional properties of beta cell subpopulations within the islet network in terms of their influence on network synchronization. The authors conclude that the most functionally coupled cell subpopulations in the islet network are not those that are most structurally coupled via gap junctions but those that are most metabolically active.
Strengths of the paper include (1) its use of an interdisciplinary collection of methods including computer simulations, FRAP to monitor functional coupling by gap junctions, the monitoring of Ca2+ oscillations in single beta cells embedded in the network, and the use of sophisticated approaches from probability theory. Most of these methods have been used and validated previously. Unfortunately, however, it was not clear what the underlying premise of the paper actually is, despite many stated intentions, nor what about it is new compared to previous studies, an additional weakness.
Although the authors state that they are trying to answer 3 critical questions, it was not clear how important these questions are in terms of significance for the field. For example, they state that a major controversy in the field is whether network structure or network function mediates functional synchronization of beta cells within the islet. However, this question is not much debated. As an example, while it is known that there can be long-range functional coupling in islets, no workers in the field believe there is a physical structure within islets that mediates this, unlike the case for CNS neurons that are known to have long projections onto other neurons. Beta cells within the islets are locally coupled via gap junctions, as stated repeatedly by the authors but these mediate short-range coupling. Thus, there are clearly functional correlations over long ranges but no structures, only correlated activity. This weakness raises questions about the overall significance of the work, especially as it seems to reiterate ideas presented previously.
We thank reviewer 4 for their positive comments, including our multidisciplinary use of mathematical models and experimental imaging techniques. We have now included an additional model of slow oscillations (the Integrated Oscillator Model) to improve our conclusions. We also thank reviewer 4 for the insightful comments. We have carefully reviewed each comment and made significant changes to the manuscript accordingly. In particular, we have significantly rewritten the introduction and discussion attempting to clarify what is new in our manuscript and what is previously shown. Additionally, we agree with the reviewers’ sentiment that there is little debate over whether, for example, there are physical structures within the islet that mediate long-range functional connections. However, there is current debate over whether functional beta-cell subpopulations can dictate islet dynamics (see [11]–[13]). This debate can be framed by observing whether these functional subpopulations emerge from the islet due to physical connections (structural network) or something more nuisance (such as intrinsic dynamics). We have reframed the introduction and discussion to clarify this debate as well as more clearly state the premise of the paper.
Specific Comments
1). The authors state it is well accepted that the disruption of gap junctional coupling is a pathophysiological characteristic of diabetes, but this is not an opinion widely accepted by the field, although it has been proposed. The authors should scale back on such generalizations, or provide more compelling evidence to support such a claim.
Thank you for pointing this out, we have provided more specific citations and changes the wording from “well accepted” to “has been documented”. See Discussion page 13 lines 415-416.
- The paper relies heavily on simulations performed using a version of the model of Cha et al (2011). While this is a reasonable model of fast bursting (e.g. oscillations having periods <1 min.), the Ca2+ oscillations that were recorded by the authors and shown in Fig. 2b of the manuscript are slow oscillations with periods of 5 min and not <1 min, which is a weakness of the model in the current context. Furthermore, the model outputs that are shown lack the well-known characteristics seen in real islets, such as fast-spiking occurring on prolonged plateaus, again as can be seen by comparing the simulated oscillations shown in Fig. 1d with those in Fig. 2b. It is recommended that the simulations be repeated using a more appropriate model of slow oscillations or at least using the model of Cha et al but employed to simulate in slower bursting.
The reviewer raises an important point and caveat associated with our simulated model and experimental data. This point was also made by other reviewers, and a similar response to this comment can be found elsewhere in response to reviewer 2 point 6. To address this comment, we have performed several additional experiments and analyses:
We collected additional Ca2+ (to identify the functional network and hubs) and FRAP data (to assess gap junction permeability) in islets which show either pure slow, pure fast, or mixed oscillations. We generated networks based on each time scale to compare with FRAP gap junction permeability data. We found that the conclusions of our first draft to be consistent across all oscillation types. There was no relationship between gap junction conductance, as approximated using FRAP, and normalized degree for slow (Figure 3j), fast (Figure 3 Supp 1d,e), or mixed (Figure 3 Supp 1g,h) oscillations. We also include discussion of these conclusions - See Results page 7 lines 184-186 and lines 188-191, Discussion page 12 lines 357-360.
We also performed additional simulations with a coupled ‘Integrated Oscillator Model’ which shows slow oscillations because of metabolic oscillations (Figure 2). We compared connectivity with gap junction coupling and underlying cell parameters. In this case, there is an association between functional and structural networks, with highly-connected hub cells showing higher gap junction conductance (Figure 2f) but also low KATP channel conductance (gKATP) (Figure 2e). However, there are some caveats to these findings – given the nature of the IOM model, we were limited to simulating smaller islets (260 cells) and less heterogeneity in the calcium traces was observed. Additional analysis suggests the greater association between functional and structural networks in this model was a result of the smaller islets, and the association was also dependent on threshold (unlike in the Cha-Noma fast oscillator model) robust. These limitations and results are discussed further (Discussion page 11 lines 344-354).
Additionally, in the IOM, the underlying cell dynamics of highly-connected hub cells are differentiated by KATP channel conductance (gKATP), which is different than in the fast oscillator model (differentiated by metabolism, kglyc). However this difference between models can be linked to differences in the way duty cycle is influenced by gKATP and kglyc (Figure 1h, Figure 2g). In each model there was a similar association between duty cycle and highly-connected hub cells. We also discuss these findings (Discussion page 11 lines 334-343).
Overall these results and discussion with respect to the coupled IOM oscillator model can be found in Figure 2, Results page 6 lines 128-156 and Discussion page 11 lines 332-354.
- Much of the data analyzed whether obtained via simulation or through experiment seems to produce very small differences in the actual numbers obtained, as can be seen in the bar graphs shown in Figs. 1e,g for example (obtained from simulations), or Fig. 2j (obtained from experimental measurements). The authors should comment as to why such small differences are often seen as a result of their analyses throughout the manuscript and why also in many cases the observed variance is high. Related to the data shown, very few dots are shown in Figs. 1eg or Fig 4e and 4h even though these points were derived from simulations where 100s of runs could be carried out and many more points obtained for plotting. These are weaknesses unless specific and convincing explanations are provided.
We thank the reviewer for these comments, which are similar to those of reviewer 2 (point 4) and reviewer 3 (point 6). Indeed there is some variability between cells in both simulations and experiments related to the metabolic activity in hubs and non-hubs. The variability points to potentially other factors being involved in determining hubs beyond simply kglyc, including a minor role for gap junction coupling structural network and potentially cell position and other intrinsic factors. We now discuss this point – see Discussion page 12 lines 364-266.
The differences between hubs and nonhubs appear small because the value of kglyc is very small. For figure 1e, the average kglyc for nonhubs was 1.26x10-4 s-1 (which is the average of the distribution because most cells are non hubs) while the average kglyc for hubs was 1.4x10-4 s-1 which is about half of a standard deviation higher. The paired t-test controls for the small value of average kglyc.
For simulation data each of the 5 dots corresponds to a simulated islet averaged over 1000 cells (or 260 cells for coupled IOM). The computational resources are high to generate such data so it is not feasible to conduct 100s of runs. Again, we note the comparisons between hubs and non-hubs are paired, and we find statistically significant differences for kglyc in figure 1 using only 5 paired data points. That we find these differences indicates the substantial difference between hubs and non-hubs. This is further supported all effect sizes being much greater than 0.8 for all significantly different findings (Cha Noma - kglyc: 2.85, gcoup: 0.82) (IOM: gKATP: 1.27, gcoup: 2.94) – We have included these effect sizes in the captions see Figure 1 and 2 captions (pages 34,
To consider all of the available data rather than the average across an entire islet, we created a kernel density estimate the kglyc for hubs and nonhubs created by concatenating every single cell in each of the five islets. A kstest results in a highly significant difference (P<0.0001) between these two distributions.
Author response image 1.
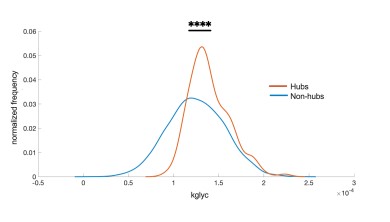
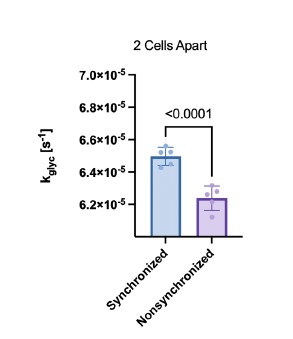
- The data shown in Fig. 4i,j are intended to compare long-range synchronization at different distances along a string of coupled cells but the difference between the synchronized and unsynchronized cells for gcoup and Kglyc was subtle, very much so.
Thank you for pointing out these subtle differences. The y-axis scale for i and j is broad to allow us to represent all distances on a single plot. After correction for multiple comparison, the differences were still statistically significant. As the reviewer mentioned in point 3, each plot contains only five data points, each of which represent the average of a single simulated islet, therefore we are not concerned about statistical significance coming from too large of a sample size. We also checked the differences between synchronized and nonsynchronized cell pairs in figure 4 panels e and h (now figure 5 e, h). These are the same data as i and j but normalized such that all of the distances could be averaged together. We again found statistical significance between synchronized and non-synchronized cell pairs. As can be seen in Author response image 2 the difference between synchronized and non-synchronized cell pairs is greater than the variability between simulated islets. Thus, in this case the variability is not substantial.
Author response image 2.
- The data shown in Fig. 5 for Cx36 knockout islets are used to assess the influence of gap junctional coupling, which is reasonable, but it would be reassuring to know that loss of this gene has no effects on the expression of other genes in the beta cell, especially genes involved with glucose metabolism.
This is an important point. Previous studies have assessed that no significant change in NAD(P)H is observed in Cx36 deficient islets – see Benninger et al J.Physiol 2011 [14]. Islet architecture is also retained. Further the insulin secretory response of dissociated Cx36 knockout beta cells is the same as that of dissociated wildtype beta cells, further indicating no significant defect in the intrinsic ability of the beta cell to release insulin – see Benninger et al J.Physiol 2011 [14]. We now Mention these findings in the discussion. See Discussion page 14 lines 459-464.
- In many places throughout the paper, it is difficult to ascertain whether what is being shown is new vs. what has been shown previously in other studies. The paper would thus benefit strongly from added text highlighting the novelty here and not just restating what is known, for instance, that islets can exhibit small-world network properties. This detracts from the strengths of the paper and further makes it difficult to wade through. Even the finding here that metabolic characteristics of the beta cells can infer profound and influential functional coupling is not new, as the authors proposed as much many years ago. Again, this makes it difficult to distill what is new compared to what is mainly just being confirmed here, albeit using different methods.
Thank you for the suggestion, we have made significant modifications throughout the Introduction, Discussion and Results to be clearer about what is known from previous work and what is newly found in this manuscript.
Reviewer #5 (Public Review):
The authors use state-of-the-art computation, experiment, and current network analysis to try and disaggregate the impact of cellular metabolism driving cellular excitability and structural electrical connections through gap junctions on islet synchronization. They perform interesting simulations with a sophisticated mathematical model and compare them with closely associated experiments. This close association is impressive and is an excellent example of using mathematics to inform experiments and experimental results. The current conclusions, however, appear beyond the results presented. The use of functional connectivity is based on correlated calcium traces but is largely without an understood biophysical mechanism. This work aims to clarify such a mechanism between metabolism and structural connection and comes out on the side of metabolism driving the functional connectivity, but both are required and more nuanced conclusions should be drawn.
We thank reviewer 5 for their positive comments, including our multifaceted experimental and computational techniques. We also found the reviewers careful reading and thoughtful comments to be very helpful and we have worked to integrate each comment into our manuscript. It is evident from the reviewer comments that we did not clearly explain what was meant by our conclusions concerning the functional network reflecting metabolism rather than gap junctions. We have conducted significant rewriting to show that we are not concluding that communication (metabolic or electric) occurs due to conduits other than gap junctions. Rather, our data suggest that the functional network (which reflects calcium synchronization) reflects intrinsic dynamics of the cells, which include metabolic rates, more than individual gap junction connections.
References referred to in this response to reviewers document:
[1] A. Stožer et al., “Functional connectivity in islets of Langerhans from mouse pancreas tissue slices,” PLoS Comput Biol, vol. 9, no. 2, p. e1002923, 2013.
[2] N. L. Farnsworth, A. Hemmati, M. Pozzoli, and R. K. Benninger, “Fluorescence recovery after photobleaching reveals regulation and distribution of connexin36 gap junction coupling within mouse islets of Langerhans,” The Journal of physiology, vol. 592, no. 20, pp. 4431–4446, 2014.
[3] C.-L. Lei, J. A. Kellard, M. Hara, J. D. Johnson, B. Rodriguez, and L. J. Briant, “Beta-cell hubs maintain Ca2+ oscillations in human and mouse islet simulations,” Islets, vol. 10, no. 4, pp. 151–167, 2018.
[4] N. R. Johnston et al., “Beta cell hubs dictate pancreatic islet responses to glucose,” Cell metabolism, vol. 24, no. 3, pp. 389–401, 2016.
[5] V. Kravets et al., “Functional architecture of pancreatic islets identifies a population of first responder cells that drive the first-phase calcium response,” PLoS Biology, vol. 20, no. 9, p. e3001761, 2022.
[6] H. Ren et al., “Pancreatic α and β cells are globally phase-locked,” Nature Communications, vol. 13, no. 1, p. 3721, 2022.
[7] A. Stožer et al., “From Isles of Königsberg to Islets of Langerhans: Examining the function of the endocrine pancreas through network science,” Frontiers in Endocrinology, vol. 13, p. 922640, 2022.
[8] J. Zmazek et al., “Assessing different temporal scales of calcium dynamics in networks of beta cell populations,” Frontiers in physiology, vol. 12, p. 337, 2021.
[9] M. E. Corezola do Amaral et al., “Caloric restriction recovers impaired β-cell-β-cell gap junction coupling, calcium oscillation coordination, and insulin secretion in prediabetic mice,” American Journal of Physiology-Endocrinology and Metabolism, vol. 319, no. 4, pp. E709–E720, 2020.
[10] J. M. Dwulet, J. K. Briggs, and R. K. P. Benninger, “Small subpopulations of beta-cells do not drive islet oscillatory [Ca2+] dynamics via gap junction communication,” PLOS Computational Biology, vol. 17, no. 5, p. e1008948, May 2021, doi: 10.1371/journal.pcbi.1008948.
[11] B. E. Peercy and A. S. Sherman, “Do oscillations in pancreatic islets require pacemaker cells?,” Journal of Biosciences, vol. 47, no. 1, pp. 1–11, 2022.
[12] G. A. Rutter, N. Ninov, V. Salem, and D. J. Hodson, “Comment on Satin et al.‘Take me to your leader’: an electrophysiological appraisal of the role of hub cells in pancreatic islets. Diabetes 2020; 69: 830–836,” Diabetes, vol. 69, no. 9, pp. e10–e11, 2020.
[13] L. S. Satin and P. Rorsman, “Response to comment on satin et al.‘Take me to your leader’: An electrophysiological appraisal of the role of hub cells in pancreatic islets. Diabetes 2020; 69: 830–836,” Diabetes, vol. 69, no. 9, pp. e12–e13, 2020.
[14] R. K. Benninger, W. S. Head, M. Zhang, L. S. Satin, and D. W. Piston, “Gap junctions and other mechanisms of cell–cell communication regulate basal insulin secretion in the pancreatic islet,” The Journal of physiology, vol. 589, no. 22, pp. 5453–5466, 2011.
[15] R. Fried, Erectile dysfunction as a cardiovascular impairment. Academic Press, 2014. [16] T. Pipatpolkai, S. Usher, P. J. Stansfeld, and F. M. Ashcroft, “New insights into KATP channel gene mutations and neonatal diabetes mellitus,” Nature Reviews Endocrinology, vol. 16, no. 7, pp. 378–393, 2020.
[17] A. M. Notary, M. J. Westacott, T. H. Hraha, M. Pozzoli, and R. K. P. Benninger, “Decreases in Gap Junction Coupling Recovers Ca2+ and Insulin Secretion in Neonatal Diabetes Mellitus, Dependent on Beta Cell Heterogeneity and Noise,” PLOS Computational Biology, vol. 12, no. 9, p. e1005116, Sep. 2016, doi: 10.1371/journal.pcbi.1005116.
[18] J. V. Rocheleau, G. M. Walker, W. S. Head, O. P. McGuinness, and D. W. Piston, “Microfluidic glucose stimulation reveals limited coordination of intracellular Ca2+ activity oscillations in pancreatic islets,” Pro ceedings of the National Academy of Sciences, vol. 101, no. 35, pp. 12899–12903, 2004. [19] R. K. Benninger, M. Zhang, W. S. Head, L. S. Satin, and D. W. Piston, “Gap junction coupling and calcium waves in the pancreatic islet,” Biophysical journal, vol. 95, no. 11, pp. 5048–5061, 2008.
-
eLife assessment
The manuscript describes a set of detailed modeling and experimental studies to disentangle the respective roles of gap junctional electrical vs. metabolic coupling mechanisms in the synchronization of islet activity. This is of interest due to the importance of islet synchronization and generally islet network properties in the regulation of insulin secretion from the pancreas. The significance of the findings was judged to be mostly important and the strength of evidence provided was judged to be mostly solid overall.
-
Reviewer #1 (Public Review):
Briggs et al use a combination of mathematical modelling and experimental validation to tease apart the contributions of metabolic and electronic coupling to the pancreatic beta cell functional network. A number of recent studies have shown the existence of functional beta cell subpopulations, some of which are difficult to fully reconcile with established electrophysiological theory. More generally, the contribution of beta cell heterogeneity (metabolism, differentiation, proliferation, activity) to islet function cannot be explained by existing combined metabolic/electrical oscillator models. The present studies are thus timely in modelling the islet electrical (structural) and functional networks. Importantly, the authors show that metabolic coupling primarily drives the islet functional network, giving …
Reviewer #1 (Public Review):
Briggs et al use a combination of mathematical modelling and experimental validation to tease apart the contributions of metabolic and electronic coupling to the pancreatic beta cell functional network. A number of recent studies have shown the existence of functional beta cell subpopulations, some of which are difficult to fully reconcile with established electrophysiological theory. More generally, the contribution of beta cell heterogeneity (metabolism, differentiation, proliferation, activity) to islet function cannot be explained by existing combined metabolic/electrical oscillator models. The present studies are thus timely in modelling the islet electrical (structural) and functional networks. Importantly, the authors show that metabolic coupling primarily drives the islet functional network, giving rise to beta cell subpopulations. The studies, however, do not diminish the critical role of electrical coupling in dictating glucose responsiveness, network extent as well as longer-range synchronization. As such, the studies show that islet structural and functional networks both act to drive islet activity, and that conclusions on the islet structural network should not be made using measures of the functional network (and vice versa).
Strengths:
- State-of-the-art multi-parameter modelling encompassing electrical and metabolic components.
- Experimental validation using advanced FRAP imaging techniques, as well as Ca2+ data from relevant gap junction KO animals.
- Well-balanced arguments that frame metabolic and electrical coupling as essential contributors to islet function.
- Likely to change how the field models functional connectivity and beta cell heterogeneity.
Weaknesses:
- Limitations of FRAP and electrophysiological gap junction measures not considered.
- Limitations of Cx36 (gap junction) KO animals not considered.
- Accuracy of citations should be improved in a few cases.
-
Reviewer #2 (Public Review):
In their present work, Briggs et al. combine biophysical simulations and experimental recordings of beta cell activity with analyses of functional network parameters to determine the role played by gap-junctional coupling, metabolism, and KATP conductance in defining the functional roles that the cells play in the functional networks, assess the structure-function relationship, and to resolve an important current open question in the field on the role of so-called hub cells in islets of Langerhans.
Combining differential equation-based simulations on 1000 coupled cells with demanding calcium, NAPDH, and FRAP imaging, as well as with advanced network analyses, and then comparing the network metrics with simulated and experimentally determined properties is an achievement in its own right and a major …
Reviewer #2 (Public Review):
In their present work, Briggs et al. combine biophysical simulations and experimental recordings of beta cell activity with analyses of functional network parameters to determine the role played by gap-junctional coupling, metabolism, and KATP conductance in defining the functional roles that the cells play in the functional networks, assess the structure-function relationship, and to resolve an important current open question in the field on the role of so-called hub cells in islets of Langerhans.
Combining differential equation-based simulations on 1000 coupled cells with demanding calcium, NAPDH, and FRAP imaging, as well as with advanced network analyses, and then comparing the network metrics with simulated and experimentally determined properties is an achievement in its own right and a major methodological strength. The findings have the potential to help resolve the issue of the importance of hub cells in beta cell networks, and the methodological pipeline and data may prove invaluable for other researchers in the community.
However, methodologically functional networks may be based on different types of calcium oscillations present in beta cells, i.e., fast oscillations produced by bursts of electrical activity, slow oscillations produced by metabolic/glycolytic oscillations, or a mixture of both. At present, the authors base the network analyses on fast oscillations only in the case of simulated traces and on a mixture of fast and slow oscillations in the case of experimental traces. Since different networks may depend on the studied beta cell properties to a different extent (e.g., fast oscillation-based networks may, more importantly, depend on electrical properties and slow oscillation-based networks may more strongly depend on metabolic properties), it is important that in drawing the conclusions the authors separately address the influence of a cell's electrical and metabolic properties on its functional role in the network based on fast oscillations, slow oscillations, or a mixture of both. -
Reviewer #3 (Public Review):
Over the past decade, novel approaches to understanding beta cell connectivity and how that contributes to the overall function of the pancreatic islet have emerged. The application of network theory to beta cell connectivity has been an extremely useful tool to understand functional hierarchies amongst beta cells within an islet. This helps to provide functional relevance to observations from structural and gene expression data that beta cells are not all identical.
There are a number of "controversies" in this field that have arisen from the mathematical and subsequent experimental identification of beta "hub" cells. These are small populations of beta cells that are very highly connected to other beta cells, as assessed by applying correlation statistics to individual beta cell calcium traces across the …
Reviewer #3 (Public Review):
Over the past decade, novel approaches to understanding beta cell connectivity and how that contributes to the overall function of the pancreatic islet have emerged. The application of network theory to beta cell connectivity has been an extremely useful tool to understand functional hierarchies amongst beta cells within an islet. This helps to provide functional relevance to observations from structural and gene expression data that beta cells are not all identical.
There are a number of "controversies" in this field that have arisen from the mathematical and subsequent experimental identification of beta "hub" cells. These are small populations of beta cells that are very highly connected to other beta cells, as assessed by applying correlation statistics to individual beta cell calcium traces across the islet.
In this paper Briggs et al set out to answer the following areas of debate:
1. They use computational datasets, based on established models of beta cells acting in concert (electrically coupled) within an islet-like structure, to show that it is similarities in metabolic parameters rather than "structural" connections (ie proximity which subserves gap junction coupling) that drives functional network behaviour. Whilst the computational models are quite relevant, the fact that the parameters (eg connectivity coefficients) are quite different to what is measured experimentally, confirm the limitations of this model. Therefore it was important for the authors to back up this finding by performing both calcium and metabolic imaging of islet beta cells. These experimental data are reported to confirm that metabolic coupling was more strongly related to functional connectivity than gap junction coupling. However, a limitation here is that the metabolic imaging data confirmed a strong link between disconnected beta cells and low metabolic coupling but did not robustly show the opposite. Similarly, I was not convinced that the FRAP studies, which indirectly measured GJ ("structural") connections were powered well enough to be related to measures of beta cell connectivity.
2. The group goes on to provide further analytical and experimental data with a model of increasing loss of GJ connectivity (by calcium imaging islets from WT, heterozygous (50% GJ loss), and homozygous (100% loss). Given the former conclusion that it was metabolic not GJ connectivity that drives small world network behaviour, it was surprising to see such a great effect on the loss of hubs in the homs. That said, the analytical approaches in this model did help the authors confirm that the loss of gap junctions does not alter the preferential existence of beta cell connectivity and confirms the important contribution of metabolic "coupling". One perhaps can therefore conclude that there are two types of network behaviour in an islet (maybe more) and the field should move towards an understanding of overlapping network communities as has been done in brain networks.Overall this is an extremely well-written paper which was a pleasure to read. This group has neatly and expertly provided both computational and experimental data to support the notion that it is metabolic but not "structural" ie GJ coupling that drives our observations of hubs and functional connectivity. However, there is still much work to do to understand whether this metabolic coupling is just a random epiphenomenon or somehow fated, the extent to which other elements of "structural" coupling - ie the presence of other endocrine cell types, the spatial distribution of paracrine hormone receptors, blood vessels and nerve terminals are also important.
-
Reviewer #4 (Public Review):
This manuscript describes a complex, highly ambitious set of modeling and experimental studies that appear designed to compare the structural and functional properties of beta cell subpopulations within the islet network in terms of their influence on network synchronization. The authors conclude that the most functionally coupled cell subpopulations in the islet network are not those that are most structurally coupled via gap junctions but those that are most metabolically active.
Strengths of the paper include (1) its use of an interdisciplinary collection of methods including computer simulations, FRAP to monitor functional coupling by gap junctions, the monitoring of Ca2+ oscillations in single beta cells embedded in the network, and the use of sophisticated approaches from probability theory. Most of …
Reviewer #4 (Public Review):
This manuscript describes a complex, highly ambitious set of modeling and experimental studies that appear designed to compare the structural and functional properties of beta cell subpopulations within the islet network in terms of their influence on network synchronization. The authors conclude that the most functionally coupled cell subpopulations in the islet network are not those that are most structurally coupled via gap junctions but those that are most metabolically active.
Strengths of the paper include (1) its use of an interdisciplinary collection of methods including computer simulations, FRAP to monitor functional coupling by gap junctions, the monitoring of Ca2+ oscillations in single beta cells embedded in the network, and the use of sophisticated approaches from probability theory. Most of these methods have been used and validated previously. Unfortunately, however, it was not clear what the underlying premise of the paper actually is, despite many stated intentions, nor what about it is new compared to previous studies, an additional weakness.
Although the authors state that they are trying to answer 3 critical questions, it was not clear how important these questions are in terms of significance for the field. For example, they state that a major controversy in the field is whether network structure or network function mediates functional synchronization of beta cells within the islet. However, this question is not much debated. As an example, while it is known that there can be long-range functional coupling in islets, no workers in the field believe there is a physical structure within islets that mediates this, unlike the case for CNS neurons that are known to have long projections onto other neurons. Beta cells within the islets are locally coupled via gap junctions, as stated repeatedly by the authors but these mediate short-range coupling. Thus, there are clearly functional correlations over long ranges but no structures, only correlated activity. This weakness raises questions about the overall significance of the work, especially as it seems to reiterate ideas presented previously.
Specific Comments
1. The authors state it is well accepted that the disruption of gap junctional coupling is a pathophysiological characteristic of diabetes, but this is not an opinion widely accepted by the field, although it has been proposed. The authors should scale back on such generalizations, or provide more compelling evidence to support such a claim.
2. The paper relies heavily on simulations performed using a version of the model of Cha et al (2011). While this is a reasonable model of fast bursting (e.g. oscillations having periods <1 min.), the Ca2+ oscillations that were recorded by the authors and shown in Fig. 2b of the manuscript are slow oscillations with periods of 5 min and not <1 min, which is a weakness of the model in the current context. Furthermore, the model outputs that are shown lack the well-known characteristics seen in real islets, such as fast-spiking occurring on prolonged plateaus, again as can be seen by comparing the simulated oscillations shown in Fig. 1d with those in Fig. 2b. It is recommended that the simulations be repeated using a more appropriate model of slow oscillations or at least using the model of Cha et al but employed to simulate in slower bursting.
3. Much of the data analyzed whether obtained via simulation or through experiment seems to produce very small differences in the actual numbers obtained, as can be seen in the bar graphs shown in Figs. 1e,g for example (obtained from simulations), or Fig. 2j (obtained from experimental measurements). The authors should comment as to why such small differences are often seen as a result of their analyses throughout the manuscript and why also in many cases the observed variance is high. Related to the data shown, very few dots are shown in Figs. 1e-g or Fig 4e and 4h even though these points were derived from simulations where 100s of runs could be carried out and many more points obtained for plotting. These are weaknesses unless specific and convincing explanations are provided.
4. The data shown in Fig. 4i,j are intended to compare long-range synchronization at different distances along a string of coupled cells but the difference between the synchronized and unsynchronized cells for gcoup and gKglyc was subtle, very much so.
5. The data shown in Fig. 5 for Cx36 knockout islets are used to assess the influence of gap junctional coupling, which is reasonable, but it would be reassuring to know that loss of this gene has no effects on the expression of other genes in the beta cell, especially genes involved with glucose metabolism.
6. In many places throughout the paper, it is difficult to ascertain whether what is being shown is new vs. what has been shown previously in other studies. The paper would thus benefit strongly from added text highlighting the novelty here and not just restating what is known, for instance, that islets can exhibit small-world network properties. This detracts from the strengths of the paper and further makes it difficult to wade through. Even the finding here that metabolic characteristics of the beta cells can infer profound and influential functional coupling is not new, as the authors proposed as much many years ago. Again, this makes it difficult to distill what is new compared to what is mainly just being confirmed here, albeit using different methods. -
Reviewer #5 (Public Review):
The authors use state-of-the-art computation, experiment, and current network analysis to try and disaggregate the impact of cellular metabolism driving cellular excitability and structural electrical connections through gap junctions on islet synchronization. They perform interesting simulations with a sophisticated mathematical model and compare them with closely associated experiments. This close association is impressive and is an excellent example of using mathematics to inform experiments and experimental results. The current conclusions, however, appear beyond the results presented. The use of functional connectivity is based on correlated calcium traces but is largely without an understood biophysical mechanism. This work aims to clarify such a mechanism between metabolism and structural connection …
Reviewer #5 (Public Review):
The authors use state-of-the-art computation, experiment, and current network analysis to try and disaggregate the impact of cellular metabolism driving cellular excitability and structural electrical connections through gap junctions on islet synchronization. They perform interesting simulations with a sophisticated mathematical model and compare them with closely associated experiments. This close association is impressive and is an excellent example of using mathematics to inform experiments and experimental results. The current conclusions, however, appear beyond the results presented. The use of functional connectivity is based on correlated calcium traces but is largely without an understood biophysical mechanism. This work aims to clarify such a mechanism between metabolism and structural connection and comes out on the side of metabolism driving the functional connectivity, but both are required and more nuanced conclusions should be drawn.
-
