Intrinsic mechanisms in the gating of resurgent Na+ currents
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
After more than 20 years of intensive research the molecular machinery of Resurgent Currents (INaR), a non-canonical identity of currents mediated by voltage-activated sodium channels is still a mystery. In this paper, Ransdell and colleagues advance the conceptual framework with new experimental insight and a new kinetic model of INaR.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
The resurgent component of the voltage-gated sodium current (I NaR ) is a depolarizing conductance, revealed on membrane hyperpolarizations following brief depolarizing voltage steps, which has been shown to contribute to regulating the firing properties of numerous neuronal cell types throughout the central and peripheral nervous systems. Although mediated by the same voltage-gated sodium (Nav) channels that underlie the transient and persistent Nav current components, the gating mechanisms that contribute to the generation of I NaR remain unclear. Here, we characterized Nav currents in mouse cerebellar Purkinje neurons, and used tailored voltage-clamp protocols to define how the voltage and the duration of the initial membrane depolarization affect the amplitudes and kinetics of I NaR . Using the acquired voltage-clamp data, we developed a novel Markov kinetic state model with parallel (fast and slow) inactivation pathways and, we show that this model reproduces the properties of the resurgent, as well as the transient and persistent, Nav currents recorded in (mouse) cerebellar Purkinje neurons. Based on the acquired experimental data and the simulations, we propose that resurgent Na + influx occurs as a result of fast inactivating Nav channels transitioning into an open/conducting state on membrane hyperpolarization, and that the decay of I NaR reflects the slow accumulation of recovered/opened Nav channels into a second, alternative and more slowly populated, inactivated state. Additional simulations reveal that extrinsic factors that affect the kinetics of fast or slow Nav channel inactivation and/or impact the relative distribution of Nav channels in the fast- and slow-inactivated states, such as the accessory Navβ4 channel subunit, can modulate the amplitude of I NaR .
Article activity feed
-

-

Author Response:
Reviewer #3 (Public Review):
INaR is related to an alternative inactivation mode of voltage activated sodium channels. It was suggested that an intracellular charged particle blocks the sodium channel alpha subunit from the intracellular space in addition to the canonical fast inactivation pathway. Putative particles revealed were sodium channel beta4 subunit and Fibroblast growth factor 14. However, abolishing the expression of neither protein does eliminate INaR. Therefore as recently suggested by several authors it is conceivable that INaR is not mediated by a particle driven mechanism at all. Instead, these and other proteins might bind to the pore forming alpha subunit and endow it with an alternative inactivation pathway as envisioned in this paper by the authors.
The main experimental findings were (1) The …
Author Response:
Reviewer #3 (Public Review):
INaR is related to an alternative inactivation mode of voltage activated sodium channels. It was suggested that an intracellular charged particle blocks the sodium channel alpha subunit from the intracellular space in addition to the canonical fast inactivation pathway. Putative particles revealed were sodium channel beta4 subunit and Fibroblast growth factor 14. However, abolishing the expression of neither protein does eliminate INaR. Therefore as recently suggested by several authors it is conceivable that INaR is not mediated by a particle driven mechanism at all. Instead, these and other proteins might bind to the pore forming alpha subunit and endow it with an alternative inactivation pathway as envisioned in this paper by the authors.
The main experimental findings were (1) The amplitude of INaR is independent of the voltage of the preceding step. (2) The peak amplitudes of INaR are dependent on the time of the depolarizing step but independent of the sodium driving force. (3) INaT and INaR are differential sensitive to recovery from inactivation. According to their experimental data the authors put forward a kinetic scheme that was fitted to their voltage-clamp patch-clamp recordings of freshly isolated Purkinje cells. The kinetic model proposed here has one open state and three inactivated states, two states related to fast inactivation (IF1, IF2) and one state related to a slower process (IS). Notably IS and IF are not linked directly in the kinetic scheme.
In my humble opinion, the proposed kinetic model fails to explain important experimental aspects and falls short to be related to the molecular machinery of sodium channels as outlined below. Still it is due time to advance the concepts of INaR. The new experimental findings of the authors are important in this respect and some ideas of the new model might be integrated in future kinetics schemes. In addition, the framework of INaR is not easy to get hold on with lots of experimental findings in the literature. Likely, my review falls also short in some aspects. Discussion is much needed and appreciated.
INaT & INaR decay The authors stated that decay speed of INaT and INaR is different and hence different mechanisms are involved. However at a given voltage (-45 mV) they have nicely illustrated (Fig. 2D and in the simulation Fig. 3H) that this is not the case. This statement is also not compatible with the used Markov model. That is because (at a given voltage) the decay of both current identities proceed from the same open state. Apparent inactivation time constants might be different, though, due to the transition to the on state.
We apologize that the language used was confusing. Our suggestion that there is more than one pathway for inactivation (from an open/conducting state) is the observation that the decay of INaT being biexponential at steady-state voltages. In the revised manuscript, we point out (lines 546-549) that, at some voltages, the slower of the two decay time constants (of INaT) is identical to the time constant of INaR decay. We also discuss how this observation was previously (Raman and Bean, 2001) interpreted.
Accumulation in the IS state after INaT inactivation in IF1 and IF2 has to proceed through closed states. How is this compatible with current NaV models? The authors have addressed this issue in the discussion. The arguments they have brought forward are not convincing for me since toxins and mutations are grossly impairing channel function.
Thank you for this comment. We would like to point out that, in our Markov model, Nav channels may accumulate in IS through either the closed state or open state. This requires, of course, that Nav channels can recover from inactivation prior to deactivation. While we agree that toxins and mutations can grossly impair channel function, we think these studies remain crucial in revealing the potential gating mechanisms of Nav channel pore-forming subunits, and how these mechanisms may vary across cell types that express different combinations of accessory proteins.
Fast inactivation - parallel inactivation pathways Related to the comment above the motivation to introduce a second fast-inactivated state IF2 is not clear. Using three states for inactivation would imply three inactivation time constants (O->IF1, IF1->IF2, O->IS) which are indeed partially visible in the simulation (Fig. 3). However, experimental data of INaT inactivation seldom require more than one time constant for fast inactivation. Importantly the authors do not provide data on INaT inactivation of the model in Fig. 3. Fast Inactivation is mapped to the binding of the IFM particle. In this model at slightly negative potential IF1 and IF2 reverse from absorbing states to dissipating states. How is this compatible with the IFM mechanism? Additionally, the statements in the discussion are not helpful, either a second time constants is required for IF (two distinct states, with two time constants) or not.
We thank this Reviewer for this comment. We tried to developed the model based on previous data on Nav channel inactivation. Indeed, much experimental data exists for the fast inactivation pathway (O -> IF1). As we noted in the discussion, without the inclusion of the IF2 state, we were unable to fully reproduce our experimental data, which led us to add the IF2 state. As with all model development, we balanced the need to faithfully reproduce the experimental data with efforts to limit the complexity of the model structure. In addition, as noted in the Methods section, our routine is an automatic parameter optimization routine that seeks to minimize the error between simulation and experiments. We can never be sure that we have found an absolute minimum, or that the optimization got stuck at a local minimum when simulating without inclusion of IF2. In other words, there may be a parameter set that sufficiently fits the data without inclusion of IF2, but we were unable to find it. As a safeguard against local minima, we used multistarts of the optimization routine with different initial parameter sets. In each case, we were unable to find a sufficiently acceptable parameter set.
We agree with this Reviewer that at slightly negative potentials (compared to strong depolarizations), channels exit the IF1 state at different rates, although we would point out that channels dissipate from the IF1 state (accumulating into IS1) under both conditions (see Figure 8B-C). This requires the binding and unbinding of the IFM motif to occur with some voltagesensitivity. We believe this to be a possibility in light of evidence that suggests IFM binding (and fast-inactivation) is an allosteric effect (Yan et al., 2017) and evidence showing that mutations in the pore-lining S6 segments can give rise to shifts of the voltage-dependence of fast inactivation without correlated shifts in the voltage-dependence of activation (Cervenka et al., 2018). However, it remains unclear how voltage-sensing in the Nav channel interact with fast- and slow-inactivation processes.
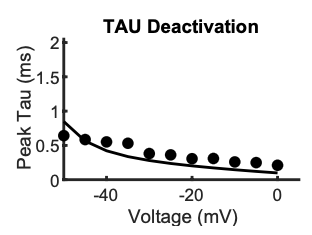
Due to space constraints in Figure 3, we did not show a plot of INaT voltage dependence. However, below, please find the experimental data (points), and simulated (line) INaT in our model.

Differential recovery of INaT & INaR Different kinetics for INaR and INaR are a very interesting finding. In my opinion, this data is not compatible with the proposed Markov model (and the authors do not provide data on the simulation). If INaT1 and INaT2 (Fig. 5 A) have the same amplitude the occupancy of the open state must be the same. I think there is no way to proceed differentially to the open state of INaR in subsequent steps unless e.g. slow inactivated states are introduced.
Thank you for bringing up this important point. The differential recovery of INaT and INaR indicates there are distinct Nav channel populations underlying the Nav currents in Purkinje neurons. We make this point on lines 632-635 of the revised manuscript. Because our Markov model is used to simulate a single channel population, we do not expect the model to reproduce the results shown in Figure 5. We have now added this point to the Discussion section on lines 637-640.
Kinetic scheme Comparison with the Raman-Bean model is a bit unfair unless the parameters are fitted to the same dataset used in this study. However, the authors have an important point in stating that this model could not reproduce all aspects of INaR. A more detailed discussion (and maybe analysis) of the states required for the models would be ideal including recent literature (e.g., J Physiol. 2020 Jan;598(2):381-40). Could the Raman-Bean model perform better if an additional inactivated state is introduced? Are alternative connections possible in the proposed model? How ambiguous is the model? Is given my statements above a second open state required? Finally, a better link of the introduced states to NaV structure-function relationship would be beneficial.
These are all excellent points. We absolutely agree; it was/is not our intention to “prove” that the Raman-Bean model does not fit our dataset (as you mention, with proper refinement of the parameters, some of the data may be well fit). In fact, qualitatively we found the Raman-Bean model quite consistent with our dataset (which is an excellent validation of both the model, and our data). It was our intention to show (in Figure 7) that there is good agreement between the Raman-Bean model and our experimental data for steady state inactivation (C), availability (D), and recovery from inactivation (E). While we find the magnitude of the resurgent current (F) to be markedly different than the Raman-Bean data, we now note this to likely be due to the large differences in the extracellular Na+ concentrations used in voltage-clamp experiments (lines 440-444). Our models, however, specifically differ in our parallel fast and slow inactivation pathways (Figure 7H). As seen in the Raman-Bean model, in response to a prolonged depolarizing holding potential, there is negligible inactivation, as the OB state remains absorbent until the channel is repolarized. This is primarily because the channel must transit through the Open state on repolarization. We find distinctly different behavior in our data. As seen in the experimental data shown in 7H, despite a prolonged depolarization, Nav channels begin to inactivate and accumulate in the slow inactivated state without prerequisite channel opening. This behavior is impossible to fit in the Raman-Bean model, given the topological constraint of the model requiring a single pathway through the open state from the OB state.
To that point, it is also unlikely that the addition of inactivated states to the Raman-Bean model would help fit this new dataset. Indeed, the Raman-Bean model contains 7 inactivated states. If there were a connection between OB ->I6, it is possible that direct inactivation (bypassing the O state) may help. Again, however, it is not our intention to discredit the Raman-Bean model, nor is it our intention to improve the Raman-Bean model. With new datasets, a fresh look at model topology was undertaken, which is how we developed our proposed model.
This Reviewer astutely points out a known limitation of Markov (state-chain) modeling; it is impossible to tell uniqueness, or ambiguity of the model (both with parameters as well as model topology). Following the results of Menon et al. 2009 (PNAS vol. 106 / #39 / 16829 – 16834), in which they used a state mutating genetic algorithm to vary topologies of a Markov model, our group (Mangold et al. 2021, PLoS Comp Bio) recently published an algorithm to distinctly enumerate all possible model structures using rooted graph theory (e.g. all possible combinations of models, rooted around a single open state). What we found (which is not entirely surprising) is that there are many model structures and parameter sets that adequately fit certain datasets (e.g., cardiac Nav channels).
Therefore, the goal is never to find the model (indeed we don’t propose that we have done so), but rather to find a model with acceptable fits to the data and then use that model to hypothesize why that model structure works, as well as to hypothesize higher dimensional dynamics. We make these points in the revised manuscript (lines 591-597).
We did not specifically explore the impact of a second open state in our modeling and simulation studies, but we would certainly agree that a model with a second open state may recapitulate the dataset.
-
Evaluation Summary:
After more than 20 years of intensive research the molecular machinery of Resurgent Currents (INaR), a non-canonical identity of currents mediated by voltage-activated sodium channels is still a mystery. In this paper, Ransdell and colleagues advance the conceptual framework with new experimental insight and a new kinetic model of INaR.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
In this manuscript, Ransdell et al study the gating mechanisms contributing to the generation of the resurgent sodium currents (INaR) in cerebellar Purkinje neurons. Using both experimental data and simulations, the authors propose that INaR is a result of fast inactivating sodium channels transitioning into an open state on membrane hyperpolarization and propose that the current decay is due to slow accumulation of recovered channels into another slower inactivated state. The data are compelling and uphold the idea that additional intrinsic gating mechanisms in NaV channels other than blocking particles can also generate INaR. The manuscript is well written supported by good quality data and analysis.
-
Reviewer #2 (Public Review):
This manuscript presents experiments on the kinetics and voltage-dependence of resurgent sodium current in cerebellar Purkinje neurons and a kinetic model for resurgent current. Much of the basic experimental characterization of resurgent current behavior repeats earlier data from Indira Raman and her colleagues, with no major differences (except for the magnitude of resurgent current relative to transient current, which is much larger here than in previous reports). There are also some new experimental protocols, notably those in Figures 5 and 8. The main point of the manuscript is a reinterpretation of the mechanistic basis of resurgent current. Raman and her colleagues suggested that the origin of resurgent current could be voltage-dependent open channel block by a blocking particle of some kind, with …
Reviewer #2 (Public Review):
This manuscript presents experiments on the kinetics and voltage-dependence of resurgent sodium current in cerebellar Purkinje neurons and a kinetic model for resurgent current. Much of the basic experimental characterization of resurgent current behavior repeats earlier data from Indira Raman and her colleagues, with no major differences (except for the magnitude of resurgent current relative to transient current, which is much larger here than in previous reports). There are also some new experimental protocols, notably those in Figures 5 and 8. The main point of the manuscript is a reinterpretation of the mechanistic basis of resurgent current. Raman and her colleagues suggested that the origin of resurgent current could be voltage-dependent open channel block by a blocking particle of some kind, with resurgent current representing flow of current through the channels when the blocking particle exits the channel on repolarization of the membrane. Here, the authors argue that that a particular kinetic model based on this idea presented by Raman and Bean (2001) does not explain some of the experimental behavior (notably decline of resurgent current during maintained depolarizations to strongly positive voltages) and present a different kinetic model that does account for this behavior, which is based on transitions of the channel between three distinct inactivated states. A key distinction is that in the Raman-Bean model, there are no direct transitions between the inactivated state corresponding to occupation by the hypothetical blocking particle and the "normal" inactivated state, so that channels have to pass through the open state in transitioning from the blocked state and the normal inactivated state, while in the new model, channels can transition between "fast" and "slow" inactivated states without opening. This correctly predicts that resurgent current declines during maintained depolarizations to +20 mV, while in the Raman-Bean model this does not occur, because channels stay in the blocked state at such positive voltages.
The work in the manuscript, and especially the suggestion that the phenomenon of resurgent current may not be tied to a blocking particle of some kind, seems to me to be interesting and significant, and at the least should stimulate further work on the molecular mechanisms involved. If the authors are correct, then searching for proteins with "blocking particle" motifs may be pointless. Personally, I would still favor a blocking particle mechanism, even after the results and arguments in this manuscript. It seems to me possible that simply allowing blocked channels to slowly inactivate with the blocking particle in the channel (which seems very plausible given that normal inactivation is now known to be an allosteric effect rather than occupancy the pore by the domain III-IV linker) could largely fix the limitations of the Raman-Bean model. Another argument in favor of the blocking mechanism is that behavior similar or identical to normal resurgent current is produced by free charged peptide pieces from the beta4 subunit, even if experiments have now shown that the beta4 subunit is not necessary for resurgent current in Purkinje neurons. However, the authors' arguments should at the least provoke new experiments - for example, it would be interesting to know whether "normal" non-resurgent channels blocked with beta4 peptide can inactivate while occupied by the blocking particle.
-
Reviewer #3 (Public Review):
INaR is related to an alternative inactivation mode of voltage activated sodium channels. It was suggested that an intracellular charged particle blocks the sodium channel alpha subunit from the intracellular space in addition to the canonical fast inactivation pathway. Putative particles revealed were sodium channel beta4 subunit and Fibroblast growth factor 14. However, abolishing the expression of neither protein does eliminate INaR. Therefore as recently suggested by several authors it is conceivable that INaR is not mediated by a particle driven mechanism at all. Instead, these and other proteins might bind to the pore forming alpha subunit and endow it with an alternative inactivation pathway as envisioned in this paper by the authors.
The main experimental findings were (1) The amplitude of INaR is …
Reviewer #3 (Public Review):
INaR is related to an alternative inactivation mode of voltage activated sodium channels. It was suggested that an intracellular charged particle blocks the sodium channel alpha subunit from the intracellular space in addition to the canonical fast inactivation pathway. Putative particles revealed were sodium channel beta4 subunit and Fibroblast growth factor 14. However, abolishing the expression of neither protein does eliminate INaR. Therefore as recently suggested by several authors it is conceivable that INaR is not mediated by a particle driven mechanism at all. Instead, these and other proteins might bind to the pore forming alpha subunit and endow it with an alternative inactivation pathway as envisioned in this paper by the authors.
The main experimental findings were (1) The amplitude of INaR is independent of the voltage of the preceding step. (2) The peak amplitudes of INaR are dependent on the time of the depolarizing step but independent of the sodium driving force. (3) INaT and INaR are differential sensitive to recovery from inactivation. According to their experimental data the authors put forward a kinetic scheme that was fitted to their voltage-clamp patch-clamp recordings of freshly isolated Purkinje cells. The kinetic model proposed here has one open state and three inactivated states, two states related to fast inactivation (IF1, IF2) and one state related to a slower process (IS). Notably IS and IF are not linked directly in the kinetic scheme.
In my humble opinion, the proposed kinetic model fails to explain important experimental aspects and falls short to be related to the molecular machinery of sodium channels as outlined below. Still it is due time to advance the concepts of INaR. The new experimental findings of the authors are important in this respect and some ideas of the new model might be integrated in future kinetics schemes. In addition, the framework of INaR is not easy to get hold on with lots of experimental findings in the literature. Likely, my review falls also short in some aspects. Discussion is much needed and appreciated.
INaT & INaR decay:
The authors stated that decay speed of INaT and INaR is different and hence different mechanisms are involved. However at a given voltage (-45 mV) they have nicely illustrated (Fig. 2D and in the simulation Fig. 3H) that this is not the case. This statement is also not compatible with the used Markov model. That is because (at a given voltage) the decay of both current identities proceed from the same open state. Apparent inactivation time constants might be different, though, due to the transition to the on state.
Accumulation in the IS state after INaT inactivation in IF1 and IF2 has to proceed through closed states. How is this compatible with current NaV models? The authors have addressed this issue in the discussion. The arguments they have brought forward are not convincing for me since toxins and mutations are grossly impairing channel function.
Fast inactivation - parallel inactivation pathways:
Related to the comment above the motivation to introduce a second fast-inactivated state IF2 is not clear. Using three states for inactivation would imply three inactivation time constants (O->IF1, IF1->IF2, O->IS) which are indeed partially visible in the simulation (Fig. 3). However, experimental data of INaT inactivation seldom require more than one time constant for fast inactivation. Importantly the authors do not provide data on INaT inactivation of the model in Fig. 3. Fast Inactivation is mapped to the binding of the IFM particle. In this model at slightly negative potential IF1 and IF2 reverse from absorbing states to dissipating states. How is this compatible with the IFM mechanism? Additionally, the statements in the discussion are not helpful, either a second time constants is required for IF (two distinct states, with two time constants) or not.
Differential recovery of INaT & INaR:
Different kinetics for INaR and INaR are a very interesting finding. In my opinion, this data is not compatible with the proposed Markov model (and the authors do not provide data on the simulation). If INaT1 and INaT2 (Fig. 5 A) have the same amplitude the occupancy of the open state must be the same. I think there is no way to proceed differentially to the open state of INaR in subsequent steps unless e.g. slow inactivated states are introduced.
Kinetic scheme:
Comparison with the Raman-Bean model is a bit unfair unless the parameters are fitted to the same dataset used in this study. However, the authors have an important point in stating that this model could not reproduce all aspects of INaR. A more detailed discussion (and maybe analysis) of the states required for the models would be ideal including recent literature (e.g., J Physiol. 2020 Jan;598(2):381-40). Could the Raman-Bean model perform better if an additional inactivated state is introduced? Are alternative connections possible in the proposed model? How ambiguous is the model? Is given my statements above a second open state required? Finally, a better link of the introduced states to NaV structure-function relationship would be beneficial.
-