Computational mechanisms for temporal integration in the anterior claustrum
Curation statements for this article:-
Curated by eLife

eLife Assessment
This work provides an important modeling-based framework for understanding the processes of temporal integration in the claustrum. These mechanisms could support a broader range of integrative brain function. The manuscript presents solid evidence for how claustrum may integrate temporal disparate signals via a novel computational phenomenon with neural dynamics evolving along neural trajectories as opposed to settling into fixed-point attractor states.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
The claustrum, with its extensive reciprocal connections to nearly all cortical regions, has long been hypothesized as a key hub for integrating diverse cognitive, sensory and motor information. However, despite its anatomical connectivity, whether and how it functionally integrates different inputs to generate coherent representations has remained unclear. Here, we developed a recurrent neural network (RNN) trained via supervised learning on behavioral metrics of delayed escape—a behavioral paradigm that requires integration of temporally separated task-relevant signals. A subset of RNN neurons exhibited dynamics similar to those of anterior claustral neurons during this behavior. These neurons formed a recurrent cluster, a structure supported by in vitro stimulation experiments in claustral brain slices. We analyzed the computational properties of this claustrum-like cluster via dimensionality reduction of population activity. The network showed nonlinear integration of temporally distributed inputs and increased synergistic information. Rather than settling into attractors, integrated information was dynamically encoded along continuously evolving neural trajectories. Notably, similar trajectory patterns associated with dynamic integration were observed in claustral recordings, suggesting the model’s biological plausibility. We propose that the anterior claustrum dynamically integrates task-relevant input signals over time and broadcasts the evolving representation to downstream brain regions capable of reading and interpreting it in a context-dependent manner.
Article activity feed
-

-
-

eLife Assessment
This work provides an important modeling-based framework for understanding the processes of temporal integration in the claustrum. These mechanisms could support a broader range of integrative brain function. The manuscript presents solid evidence for how claustrum may integrate temporal disparate signals via a novel computational phenomenon with neural dynamics evolving along neural trajectories as opposed to settling into fixed-point attractor states.
-
Reviewer #1 (Public review):
Summary:
In this manuscript, the authors investigate how the anterior claustrum may integrate temporally separated task-relevant signals to guide behavior in a delayed escape paradigm. Because in vivo neural recordings from claustrum during this task are extremely limited-comprising single-trial data with small neuronal samples-the authors adopt a modeling-driven approach. They train recurrent neural networks (RNNs) using only behavioral data (escape latency) to reproduce task performance and then analyze the internal dynamics of the trained networks. Within these networks, they identify a subset of units whose activity exhibits persistent responses and strong correlations with behavior, which the authors label as "claustrum-like." Using dimensionality reduction, decoding, and information-theoretic analyses, …
Reviewer #1 (Public review):
Summary:
In this manuscript, the authors investigate how the anterior claustrum may integrate temporally separated task-relevant signals to guide behavior in a delayed escape paradigm. Because in vivo neural recordings from claustrum during this task are extremely limited-comprising single-trial data with small neuronal samples-the authors adopt a modeling-driven approach. They train recurrent neural networks (RNNs) using only behavioral data (escape latency) to reproduce task performance and then analyze the internal dynamics of the trained networks. Within these networks, they identify a subset of units whose activity exhibits persistent responses and strong correlations with behavior, which the authors label as "claustrum-like." Using dimensionality reduction, decoding, and information-theoretic analyses, they argue that these units dynamically integrate conditioned stimulus (CS) and door-opening signals via nonlinear, trajectory-based population dynamics rather than fixed-point attractor states.
To bridge model predictions and biology, the authors complement the modeling with in vitro slice experiments demonstrating recurrent excitatory connectivity and prolonged activity in the anterior claustrum that depends on glutamatergic transmission. They further compare latent neural trajectories derived from previously published in vivo claustrum recordings to those observed in the RNN, reporting qualitative similarities. Based on these results, the authors propose that the claustrum implements temporal signal integration through recurrent excitatory circuitry and dynamic population trajectories, potentially supporting broader theories of integrative brain function.
Strengths:
This study addresses an important and challenging problem: how to infer population-level computation in a brain structure for which in vivo data are sparse and experimentally constrained. The authors are commendably transparent about these limitations and seek to overcome them through a principled modeling framework. The integration of behavioral modeling, RNN analysis, and slice electrophysiology is ambitious and technically sophisticated.
Several aspects stand out as strengths. First, the behavioral RNN is carefully trained and interrogated using a rich set of modern analytical tools, including cross-temporal decoding, trajectory analysis, and partial information decomposition, providing multiple complementary views of network dynamics. Second, the slice experiments convincingly demonstrate recurrent excitatory connectivity in anterior claustrum, lending biological plausibility to the model's reliance on recurrent dynamics. Third, the manuscript is clearly written, logically organized, and conceptually engaging, and it offers a coherent mechanistic hypothesis that could guide future large-scale recording experiments.
Importantly, the work has significant heuristic value: rather than merely fitting data, it attempts to generate testable computational ideas about claustral function in a regime where direct empirical access is currently limited.
Weaknesses:
Despite these strengths, the manuscript suffers from a recurring and substantial conceptual issue: systematic over-interpretation of model-data correspondence. While the modeling results are potentially insightful, the extent to which they are presented as recapitulating real claustral neural mechanisms goes beyond what the available data can support.
A fundamental limitation is that the RNN is trained solely on behavioral output, without being constrained by neural data at either single-unit or population levels. As a result, the internal network dynamics are underdetermined and non-unique. Many distinct internal solutions could plausibly generate identical behavior. However, the manuscript frequently treats the specific internal solution discovered in the RNN as if it were a close approximation of the actual claustrum circuit.
This issue is compounded by the sparse nature of the in vivo data used for comparison. The GPFA-based trajectory analyses rely on pseudo-populations and single-trial recordings, yet are interpreted as evidence for robust population-level dynamics. Because neurons were not recorded simultaneously, the inferred trajectories necessarily lack true population covariance and shared trial-to-trial variability, limiting their interpretability as genuine population dynamics. Similarly, conclusions about trajectory-based versus attractor-based computation are drawn almost exclusively from model analyses and then generalized to the biological system.
Overall, while the modeling framework is appropriate as a hypothesis-generating tool, the manuscript repeatedly crosses the line from proposing plausible mechanisms to asserting explanatory or even causal equivalence between the model and the brain. This undermines the otherwise strong contributions of the work.
Below are several specific points that warrant further clarification or revision:
(1) Tone of model-data correspondence
Numerous statements describe the RNN as "closely mimicking," "recapitulating," or being "nearly identical" to claustral neural dynamics, sometimes extending to claims about causal relationships between neural activity and behavior. Given that neural data were not used to train the model, and that only a small subset of trained networks showed the reported dynamics, these statements should be substantially softened throughout the manuscript. The RNN should be framed as providing one possible computational realization consistent with existing data, not as a close instantiation of the biological circuit.
(2) Non-uniqueness of RNN solutions
The fact that only a small fraction of trained networks exhibited "claustrum-like" clusters deserves deeper discussion. This observation raises the possibility that the identified solution is fragile or highly specific rather than canonical. The authors should explicitly discuss the non-uniqueness of internal solutions in behavior-trained RNNs, including the range of alternative network dynamics that can reproduce the same behavior. In particular, it should be clarified why the specific network exhibiting "claustrum-like" clusters is informative about claustral computation, rather than representing one arbitrary solution among many.
(3) GPFA trajectory comparisons
The qualitative similarity between RNN trajectories and GPFA-derived trajectories from sparse in vivo data is interesting but insufficient to support claims of robustness or population-level structure. Statements suggesting that these patterns are unlikely to arise from noise or random fluctuations are not justified given the single-trial, pseudo-population nature of the data. Either additional quantitative controls should be added, or the interpretation should be substantially tempered.
(4) Scope of functional claims
The discussion connecting the findings to broad theories of claustral function, global workspace, or consciousness extends well beyond the data presented. These speculative links should be clearly labeled as such and significantly reduced in strength and prominence.
The manuscript repeatedly describes the delayed escape task as an "inference-based behavioral paradigm" and states that animals "infer that a value-neutral alternative space is likely to be safer" when the CS is presented in a novel environment. While I appreciate that the US-CS association was established in a different context and that the CS is then presented in a new environment, I am not convinced that the current behavioral evidence uniquely supports an inference interpretation.
First, it is not clear that this task is widely recognized in the literature as a canonical inference task, in the sense of, for example, sensory preconditioning, transitive inference, or model-based inference paradigms. Rather, the observed effect-that CS animals escape faster to a neutral compartment than neutral-CS controls-can be parsimoniously interpreted in terms of generalized threat value, heightened fear/anxiety, or a bias toward avoidance/escape under elevated threat, without requiring an explicit inferential step about the specific safety of the alternative compartment. The fact that no prior training is needed is compatible with flexible generalization, but does not by itself demonstrate inference in a more formal computational sense.
Second, the inference claim becomes central to the manuscript's conceptual framing (e.g., the idea that rsCla supports "inference-based escape"), yet the behavioral analyses presented here and in the cited prior work do not clearly rule out simpler accounts. Clarifying this distinction would help avoid overstating both the inferential nature of the behavior and the specific role of rsCla and the RNN's "claustrum-like" cluster in supporting inference per se, as opposed to more general integration of threat-related signals with an opportunity for escape.
This manuscript presents an interesting and potentially valuable modeling-based framework for thinking about temporal integration in the claustrum, supported by solid slice physiology. However, in its current form, it overstates the degree to which the proposed RNN dynamics reflect actual claustral neural mechanisms. With substantial revision-especially a more cautious interpretation of model-data similarity and a clearer articulation of modeling limitations-the study could make a meaningful contribution as a hypothesis-generating work rather than a definitive mechanistic account.
Comments on revisions:
The authors have carefully addressed the concerns raised in the initial review. In particular, the manuscript has been substantially improved in terms of tone, conceptual clarity, and the interpretation of the modeling results. The revised version now presents a well-balanced and appropriately framed account of the work.
The study offers a compelling and useful hypothesis-generating framework for understanding temporal integration in the claustrum, and I support its publication. As a minor point, given the acknowledged limitations of pseudo-population and single-trial data, it would be preferable to slightly soften a few remaining statements that describe trajectory structure as directly "reflecting" population-level dynamics (e.g., using "consistent with" instead).
-
Reviewer #2 (Public review):
This manuscript reports the behavior of a computational model of rat claustral neurons during the performance of a behavioral task known as the delayed escape task (in this reviewer's understanding, this behavioral task was created and implemented by this group only). These authors have argued in a prior manuscript (Han et al.) that a group of neurons located "rostral to striatum" are part of the claustrum. The group names the region the "rostral to striatum claustrum." Additionally, in the Han et al. paper, the authors argue that these cells are responsible for maintaining a signal that lasts through the delay period.
The main findings of the current paper are:
(1) The authors have built a model network that was trained to show firing similar to what was reported for rats in their prior paper.
(2) The …
Reviewer #2 (Public review):
This manuscript reports the behavior of a computational model of rat claustral neurons during the performance of a behavioral task known as the delayed escape task (in this reviewer's understanding, this behavioral task was created and implemented by this group only). These authors have argued in a prior manuscript (Han et al.) that a group of neurons located "rostral to striatum" are part of the claustrum. The group names the region the "rostral to striatum claustrum." Additionally, in the Han et al. paper, the authors argue that these cells are responsible for maintaining a signal that lasts through the delay period.
The main findings of the current paper are:
(1) The authors have built a model network that was trained to show firing similar to what was reported for rats in their prior paper.
(2) The authors' analysis of model behavior is used to suggest that the model network recapitulates biological activity, including the existence of a cluster of cells mainly responsible for the delay period firing.
(3) The authors offer evidence from patch clamp recordings for excitatory interconnections among claustral neurons that are an essential feature of the model network.
A major value of the computational network is that "trials" of the network can be performed. In experiments on animals, only single trials can be used.
Concerns:
(1) This paper is based on behavioral results and neural recordings from their prior paper (Han et al.), but data, e.g. in figure 1, are not clearly identified as new or as coming from that source. Figure 1A, for example, appears to be taken directly from Han et al. No methods are given in this manuscript for the behavioral testing or the in vivo electrophysiology.
(2) Many other details are unclear. Examples include model training, the weight matrices and how these changed with training (p. 13), the equations 2 and 3 (p. 13), the sources for the constants in the equations (p. 14), the methods (anesthesia, stereotaxic coordinates, injection specifics and details for "sparse expression") for the ChrimsonR injections.
(3) The explorations of model behavior are a catalog of everything tried rather than an organized demonstration of what the model can and cannot do. The figures could be reduced in number to emphasize the key comparisons of the different clusters and the model's behavior under different conditions intended to "test" the model.
(4) On page 6, the E-E connectivity is argued from Shelton et al. (2025) and against Kim et al. (2016), but ignores Orman (2015), which to this reviewer's knowledge was the first to demonstrate such connectivity, including the long duration events and impact of planes of section.
(5) Whereas the authors are entitled to their own opinion of prior work (references 3-8), it is inappropriate to misrepresent prior work as only demonstrating a "limited function" of claustum. Additional papers by Mathur's group and Citri's group are ignored.
In summary, the authors have made a computational model that recapitulates the firing of a subset of potentially claustral neurons during a particular behavioral task (delayed escape is certainly not the only behavior that involves claustrum - see e.g., attention, salience, sleep). If the conclusion is that excitatory claustral cells must be connected to other excitatory claustral cells, such a conclusion is not new and the electrophysiological E-E metrics are not well quantified (e.g., connectivity frequency, strength of connection). If the model is intended to predict how claustrum might accomplish any other task, there is insufficient detail to evaluate the model beyond the evidence that the model creates a subset of cells that can sustain firing during the delay period in the delayed escape task.
All relevant work must be appropriately cited throughout the manuscript.
Comments on revisions:
The authors have adequately addressed the concerns that were raised in response to the first version of the manuscript.
-
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
We thank the reviewer for their constructive and insightful comments and agree with the importance of the points raised. We recognize that aspects of our original presentation may have been unclear or overly strong in their interpretation. We have therefore revised the manuscript to clarify our intended scope, moderate our claims, and strengthen the analysis. In the second paragraph of the Discussion, we have explicitly acknowledged the concerns raised by the reviewer and outlined how they have been addressed in the revised manuscript. Our detailed responses are provided below.
(1) Tone of model-data correspondence
Numerous statements describe the RNN as "closely mimicking," "recapitulating," or being …
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
We thank the reviewer for their constructive and insightful comments and agree with the importance of the points raised. We recognize that aspects of our original presentation may have been unclear or overly strong in their interpretation. We have therefore revised the manuscript to clarify our intended scope, moderate our claims, and strengthen the analysis. In the second paragraph of the Discussion, we have explicitly acknowledged the concerns raised by the reviewer and outlined how they have been addressed in the revised manuscript. Our detailed responses are provided below.
(1) Tone of model-data correspondence
Numerous statements describe the RNN as "closely mimicking," "recapitulating," or being "nearly identical" to claustral neural dynamics, sometimes extending to claims about causal relationships between neural activity and behavior. Given that neural data were not used to train the model, and that only a small subset of trained networks showed the reported dynamics, these statements should be substantially softened throughout the manuscript. The RNN should be framed as providing one possible computational realization consistent with existing data, not as a close instantiation of the biological circuit
We agree with the reviewer’s comment. The expressions noted by the reviewer (e.g., closely mimicked, nearly identical, recapitulate) will be replaced with alternative wording that conveys a more moderate meaning (Line 16-17, 65-66, 83, 96, 120, 212).
(2) Non-uniqueness of RNN solutions
The fact that only a small fraction of trained networks exhibited "claustrum-like" clusters deserves deeper discussion. This observation raises the possibility that the identified solution is fragile or highly specific rather than canonical. The authors should explicitly discuss the non-uniqueness of internal solutions in behavior-trained RNNs, including the range of alternative network dynamics that can reproduce the same behavior. In particular, it should be clarified why the specific network exhibiting "claustrum-like" clusters is informative about claustral computation, rather than representing one arbitrary solution among many.
As the reviewer pointed out, behaviorally trained RNNs can admit multiple internal solutions that produce the same behavioral output, and we acknowledge the non-uniqueness of such internal solutions. However, we do not interpret the fact that only a subset of trained RNNs exhibit dynamics similar to those observed in the claustrum as evidence that this solution is fragile. Notably, the claustrum-like dynamics emerged spontaneously during training and were not explicitly enforced. Furthermore, our finding suggests that the emergence of this particular dynamical regime depends on relatively specific structural constraints.
Our criterion for selecting RNNs that could inform the computational principles of the claustrum was their ability to reproduce the behavioral and physiological observations obtained in the delayed escape experiments. RNNs that were excluded may reflect information-processing strategies used by other brain regions or may rely on artificial logical structures. The computational demand of the task, which integrates temporally separated signals, naturally drives convergence toward networks with recurrent excitatory connectivity capable of maintaining persistent activity. Indeed, all networks that exhibited a claustrum-like cluster shared a common structural feature: strong recurrent excitatory connectivity within Cluster 1. This property is consistent with biological characteristics observed in the slice experiments shown in Fig 2.
Importantly, the computational principles derived from this RNN were found to be quantitatively consistent with in vivo single-neuron activity patterns. Specifically, analysis using an eigenvalue-based metric (λ3/Σλ) revealed the same directional effect in both the RNN and the claustrum neuron data. In addition, a leave-one-neuron-out analysis showed that this pattern was broadly distributed across in vivo claustral neurons rather than being driven by a small subset (see Fig. 4).
Taken together, these convergent lines of evidence suggest that the computational model is not simply one arbitrary solution among many possible alternatives, but rather implements a computational principle that may underlie claustral functions.
(3) GPFA trajectory comparisons
The qualitative similarity between RNN trajectories and GPFA-derived trajectories from sparse in vivo data is interesting but insufficient to support claims of robustness or population-level structure. Statements suggesting that these patterns are unlikely to arise from noise or random fluctuations are not justified, given the single-trial, pseudo-population nature of the data. Either additional quantitative controls should be added, or the interpretation should be substantially tempered.
As the reviewer pointed out, the GPFA trajectory comparison presented in the original manuscript remained largely qualitative, and we agree that this alone was insufficient to establish robustness or provide convincing evidence for population-level structure. In the revised manuscript, we have therefore added the requested quantitative analysis (see Fig. 4).
Before describing the analysis, we would like to clarify several methodological limitations associated with pseudopopulation and single-trial data. GPFA estimates latent trajectories based on assumptions about covariance structure among neurons and temporal smoothness. In pseudopopulation datasets, the true simultaneously recorded covariance structure cannot be fully reconstructed, which is an inherent limitation. Because our dataset is based on single trials, the analysis does not directly exploit trial-to-trial variability. Nevertheless, the estimation of the latent space still depends on the covariance structure among real claustral neurons, suggesting that the inferred trajectories remain tied to biologically meaningful population dynamics.
Accordingly, the quantitative metric we introduce is not entirely independent of the GPFA estimation step. Rather, it is intended to evaluate the geometric structure of the single-trial latent trajectories estimated by GPFA. We acknowledged this limitation in the revised manuscript.
Specifically, for the biological data, we reanalyzed the GPFA-derived latent trajectories in PCA space and computed an eigenvalue-based metric (λ3/Σλ). For each of the 20 time bins, we applied a sliding window of 10 bins and calculated the covariance matrix within that window. The eigenvalues of PC1, PC2, and PC3 were then obtained, and the third eigenvalue (λ3) was normalized by the total variance (Σλ = λ1 + λ2 + λ3). This metric quantifies the degree to which the trajectory locally deviates from a planar structure that can be explained by two dominant axes. An increase in λ3/Σλ indicates that the population-state trajectory forms a higher-dimensional geometric structure beyond a simple two-dimensional combination.
For the RNN data, in contrast, the activity of all units can be observed simultaneously and sufficient trial repetitions are available. Therefore, GPFA was not applied; instead, PCA was performed directly on the population activity for each trial. We then computed an average trajectory across trials and applied the same λ3/Σλ metric. Thus, although the initial dimensionality reduction steps differ between the two systems, the definition and calculation of the final quantitative metric are identical. The focus of the comparison is therefore not the dimensionality reduction technique itself, but the geometric dimensional structure of the population trajectories evolving over time.
Importantly, within the biological dataset, the GPFA estimation procedure, preprocessing steps, pseudopopulation construction, subsampling strategy, temporal alignment criteria, and smoothing parameters were applied identically across conditions. Likewise, the same analysis pipeline was used for all conditions in the RNN. If structural biases had been introduced during covariance estimation or dimensionality reduction, they would be expected to affect all conditions within each system similarly. Nevertheless, the λ3/Σλ value was consistently and significantly higher in the CS condition than in the Neutral condition, and this directional pattern was observed in both the RNN and the claustral neuron data. This suggests that the effect reflects condition-specific differences in population dynamical structure rather than artifacts arising from a particular dimensionality reduction method.
To further test whether the observed effect might be driven by a small subset of neurons or specific neuron combinations, we performed a leave-one-neuron-out analysis on the claustrum dataset. Recomputing λ3/Σλ while removing one neuron at a time showed that, in the CS group, most neurons contributed relatively evenly to this metric, whereas the Neutral group did not show such a distributed contribution pattern. This indicates that the observed three-dimensional structure is not driven by a few outlier neurons or incidental covariance patterns, but rather reflects an organized population-level phenomenon.
If the result were primarily due to structural artifacts introduced by the pseudopopulation construction or dimensionality reduction procedures, it would be unlikely for consistent selective differences to repeatedly emerge between conditions under identical analysis pipelines. The consistently higher λ3/Σλ values observed in the CS condition therefore provide indirect support that this pattern reflects condition-specific population dynamics rather than estimation bias.
Taken together, these results suggest that the observed three-dimensional structure reflects condition-specific population dynamics rather than analysis artifacts. The fact that the same quantitative metric yields consistent effects in both the RNN and claustral data further strengthens the correspondence between the two systems.
(4) Scope of functional claims
The discussion connecting the findings to broad theories of claustral function, global workspace, or consciousness extends well beyond the data presented. These speculative links should be clearly labeled as such and significantly reduced in strength and prominence.
We agree with the reviewer and stated that references to these theories are speculative, while substantially reducing both their emphasis and prominence in the manuscript (Line 444-446, 451).
(5) Comment on Conceptual Interpretation of the Behavioral Paradigm:
The manuscript repeatedly describes the delayed escape task as an "inference-based behavioral paradigm" and states that animals "infer that a value-neutral alternative space is likely to be safer" when the CS is presented in a novel environment. While I appreciate that the US-CS association was established in a different context and that the CS is then presented in a new environment, I am not convinced that the current behavioral evidence uniquely supports an inference interpretation.
First, it is not clear that this task is widely recognized in the literature as a canonical inference task, in the sense of, for example, sensory preconditioning, transitive inference, or model-based inference paradigms. Rather, the observed effect-that CS animals escape faster to a neutral compartment than neutral-CS controls-can be parsimoniously interpreted in terms of generalized threat value, heightened fear/anxiety, or a bias toward avoidance/escape under elevated threat, without requiring an explicit inferential step about the specific safety of the alternative compartment. The fact that no prior training is needed is compatible with flexible generalization, but does not by itself demonstrate inference in a more formal computational sense.
Second, the inference claim becomes central to the manuscript's conceptual framing (e.g., the idea that rsCla supports "inference-based escape"), yet the behavioral analyses presented here and in the cited prior work do not clearly rule out simpler accounts. Clarifying this distinction would help avoid overstating both the inferential nature of the behavior and the specific role of rsCla and the RNN's "claustrum-like" cluster in supporting inference per se, as opposed to more general integration of threat-related signals with an opportunity for escape.
We agree with the reviewer’s concern. First, we referred to the delayed escape behavioral task as “a behavioral paradigm that requires integration of temporally separated task-relevant signals.” (Line 7-8). We also removed references to the term inference throughout the manuscript (Line 46, 51, 67, 397).
Reviewer #2 (Public review):
We sincerely thank the reviewer for their constructive and insightful comments. Through the revision process, the manuscript has been substantially improved, with increased reproducibility, more appropriate acknowledgment of prior work, and a clearer and more logical presentation of the study.
(1) This paper is based on behavioral results and neural recordings from their prior paper (Han et al.), but data, e.g., in Figure 1, are not clearly identified as new or as coming from that source. Figure 1A, for example, appears to be taken directly from Han et al. No methods are given in this manuscript for the behavioral testing or the in vivo electrophysiology.
We agree with the reviewer that this distinction should be made clearer. In the original manuscript, we indicated in the Figure 1 legend that panels A, D, E, F, and L (left) were reproduced from Han et al. (2024). To further clarify this point, we explicitly noted this distinction again in the main text (Line 74, 85). In addition, we described the behavioral experiments and in vivo electrophysiological recordings performed in Han et al. (2024) in the Methods section and include the appropriate citation (Line 463-530).
(2) Many other details are unclear. Examples include model training, the weight matrices and how these changed with training (p. 13), equations 2 and 3 (p. 13), the sources for the constants in the equations (p. 14), the methods (anesthesia, stereotaxic coordinates, injection specifics and details for "sparse expression") for the ChrimsonR injections.
We agree with the reviewer’s comment and have revised the manuscript to provide a more detailed description of the model training procedure, weight initialization, and parameter selection.
We expanded the explanation of the model training procedure and weight initialization. Specifically, the recurrent (Wrec) and output (Wout) weight matrices were initialized using a Glorot normal distribution with a standard deviation of
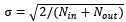 to ensure stable signal propagation during early training. In addition, we now explicitly describe the training algorithm and optimization procedure. The network was trained using the Adam optimizer implemented in TensorFlow (v2.1.0) with a batch size of 256 for 1.2 million training iterations, minimizing the per-trial loss function defined in the manuscript. We also explicitly stated how Dale’s principle was maintained throughout training: rows in W_out corresponding to inhibitory units were zeroed out, and recurrent weights were continuously constrained so that excitatory and inhibitory neurons preserved their respective positive and negative synaptic projections. To illustrate how the weight structure evolved during training, we explicitly reference Figure 2A, which visualizes the final mean inter-cluster synaptic weights and highlights the strong recurrent connectivity that emerged within Cluster 1. Regarding Equations 2 and 3 and their constants, we clarified that the target escape times used to anchor the network were based on experimentally measured behavioral latencies (48.7 s for the CS-present condition and 111.3 s for the CS-absent condition). Furthermore, the regularization coefficients (λ = 0.01 and λFR = 0.95) were selected through a grid search procedure to maintain biologically plausible firing rates while preventing overfitting.
to ensure stable signal propagation during early training. In addition, we now explicitly describe the training algorithm and optimization procedure. The network was trained using the Adam optimizer implemented in TensorFlow (v2.1.0) with a batch size of 256 for 1.2 million training iterations, minimizing the per-trial loss function defined in the manuscript. We also explicitly stated how Dale’s principle was maintained throughout training: rows in W_out corresponding to inhibitory units were zeroed out, and recurrent weights were continuously constrained so that excitatory and inhibitory neurons preserved their respective positive and negative synaptic projections. To illustrate how the weight structure evolved during training, we explicitly reference Figure 2A, which visualizes the final mean inter-cluster synaptic weights and highlights the strong recurrent connectivity that emerged within Cluster 1. Regarding Equations 2 and 3 and their constants, we clarified that the target escape times used to anchor the network were based on experimentally measured behavioral latencies (48.7 s for the CS-present condition and 111.3 s for the CS-absent condition). Furthermore, the regularization coefficients (λ = 0.01 and λFR = 0.95) were selected through a grid search procedure to maintain biologically plausible firing rates while preventing overfitting.We detailed the surgical procedures that were previously omitted. This includes the specific anesthesia protocol (sodium pentobarbital, 50 mg/kg, i.p.), stereotaxic mounting, and the exact coordinates for the rsCla (AP +2.95, ML ±1.95, DV -3.85 mm). To define "sparse expression," we specified that the AAV was diluted 1:4 in sterile saline. Finally, we included the precise injection parameters: delivery at 20 nL/min via a pressure injection system, with the pipette left in place for 10 minutes post-infusion to ensure adequate diffusion. (Line 635, 636-639, 641-643). We have added these contents in the Methods section.
(3) The explorations of model behavior are a catalog of everything tried rather than an organized demonstration of what the model can and cannot do. The figures could be reduced in number to emphasize the key comparisons of the different clusters and the model's behavior under different conditions, intended to "test" the model.
We agree with the reviewer’s comment and have reorganized the figures to focus on the key results. Specifically, we separated the original figures so that they correspond to (1) Presentation of an RNN model consistent with the results of actual claustral recordings, (2) identification of dimensionality-reduced population activity patterns in the model, (3) comparison of these patterns with population activity patterns derived from recorded claustral neurons, (4) proposal of a nonlinear integration mechanism, and (5) the suggestion that such integration may be implemented through dynamic coding. Using this figure organization, we first identify RNN models trained on behavioral metrics whose dynamics are consistent with experimental claustral recordings. We then compare the dimensionality-reduced population activity patterns of these models with those derived from recorded claustral neurons to evaluate their biological plausibility. After selecting the models that satisfy this criterion, we perform further analyses that would be difficult to achieve using real neural recordings alone. These analyses ultimately allow us to propose dynamic coding exhibiting nonlinear integration as a plausible computational mechanism.
(4) On page 6, the E-E connectivity is argued from Shelton et al. (2025) and against Kim et al. (2016), but ignores Orman (2015), which, to this reviewer's knowledge, was the first to demonstrate such connectivity, including the long-duration events and impact of planes of section.
We agree with the reviewer’s suggestion and will include a reference to Orman (2015). We have clarified that neuronal activity can persist for extended periods and that such persistent activity has been observed in claustral slices prepared at a specific slicing angle (Line 144).
(5) Whereas the authors are entitled to their own opinion of prior work (references 3-8), it is inappropriate to misrepresent prior work as only demonstrating a "limited function" of claustrum. Additional papers by Mathur's group and Citri's group are ignored.
We agree with the reviewer’s comment and have revised the relevant sentences in the Introduction section. We also included and acknowledged the contributions of previous studies by the Mathur group and the Citri group by adding additional references to their works (Line 36, 429).
In summary, the authors have made a computational model that recapitulates the firing of a subset of potentially claustral neurons during a particular behavioral task (delayed escape is certainly not the only behavior that involves claustrum - see e.g., attention, salience, sleep). If the conclusion is that excitatory claustral cells must be connected to other excitatory claustral cells, such a conclusion is not new, and the electrophysiological E-E metrics are not well quantified (e.g., connectivity frequency, strength of connection). If the model is intended to predict how the claustrum might accomplish any other task, there is insufficient detail to evaluate the model beyond the evidence that the model creates a subset of cells that can sustain firing during the delay period in the delayed escape task.
All relevant work must be appropriately cited throughout the manuscript.
Regarding the E–E metric, we obtained the following result. When including recordings in which the whole-cell recording could not be completed, optogenetically evoked responses were observed in 38 out of 43 patched cells. This suggests that approximately 90% of the cells receive intra-claustral excitatory input. However, the current dataset does not allow us to quantify the connection probability or the strength of these connections.
As the reviewer pointed out, the RNN developed in this study is specifically designed for the delayed escape task, and we do not intend to claim direct generalization to other proposed functions of the claustrum, such as attention, salience, or sleep. The goal of this study is to computationally characterize the temporal integration mechanism of the claustrum observed in this specific task. We have included this in the Discussion section. In the second paragraph of the Discussion, we have explicitly acknowledged the concerns raised by the reviewer and outlined how they have been addressed in the revised manuscript.
-
-
eLife Assessment
This work provides an important modeling-based framework for understanding the processes of temporal integration in the claustrum. These mechanisms could support a broader range of integrative brain function. However, at present, the evidence remains at least in part incomplete, primarily because of over-interpretation of the results and their connection to neurophysiology.
-
Reviewer #1 (Public review):
Summary:
In this manuscript, the authors investigate how the anterior claustrum may integrate temporally separated task-relevant signals to guide behavior in a delayed escape paradigm. Because in vivo neural recordings from claustrum during this task are extremely limited - comprising single-trial data with small neuronal samples - the authors adopt a modeling-driven approach. They train recurrent neural networks (RNNs) using only behavioral data (escape latency) to reproduce task performance and then analyze the internal dynamics of the trained networks. Within these networks, they identify a subset of units whose activity exhibits persistent responses and strong correlations with behavior, which the authors label as "claustrum-like." Using dimensionality reduction, decoding, and information-theoretic …
Reviewer #1 (Public review):
Summary:
In this manuscript, the authors investigate how the anterior claustrum may integrate temporally separated task-relevant signals to guide behavior in a delayed escape paradigm. Because in vivo neural recordings from claustrum during this task are extremely limited - comprising single-trial data with small neuronal samples - the authors adopt a modeling-driven approach. They train recurrent neural networks (RNNs) using only behavioral data (escape latency) to reproduce task performance and then analyze the internal dynamics of the trained networks. Within these networks, they identify a subset of units whose activity exhibits persistent responses and strong correlations with behavior, which the authors label as "claustrum-like." Using dimensionality reduction, decoding, and information-theoretic analyses, they argue that these units dynamically integrate conditioned stimulus (CS) and door-opening signals via nonlinear, trajectory-based population dynamics rather than fixed-point attractor states.
To bridge model predictions and biology, the authors complement the modeling with in vitro slice experiments demonstrating recurrent excitatory connectivity and prolonged activity in the anterior claustrum that depends on glutamatergic transmission. They further compare latent neural trajectories derived from previously published in vivo claustrum recordings to those observed in the RNN, reporting qualitative similarities. Based on these results, the authors propose that the claustrum implements temporal signal integration through recurrent excitatory circuitry and dynamic population trajectories, potentially supporting broader theories of integrative brain function.
Strengths:
This study addresses an important and challenging problem: how to infer population-level computation in a brain structure for which in vivo data are sparse and experimentally constrained. The authors are commendably transparent about these limitations and seek to overcome them through a principled modeling framework. The integration of behavioral modeling, RNN analysis, and slice electrophysiology is ambitious and technically sophisticated.
Several aspects stand out as strengths. First, the behavioral RNN is carefully trained and interrogated using a rich set of modern analytical tools, including cross-temporal decoding, trajectory analysis, and partial information decomposition, providing multiple complementary views of network dynamics. Second, the slice experiments convincingly demonstrate recurrent excitatory connectivity in the anterior claustrum, lending biological plausibility to the model's reliance on recurrent dynamics. Third, the manuscript is clearly written, logically organized, and conceptually engaging, and it offers a coherent mechanistic hypothesis that could guide future large-scale recording experiments.
Importantly, the work has significant heuristic value: rather than merely fitting data, it attempts to generate testable computational ideas about claustral function in a regime where direct empirical access is currently limited.
Weaknesses:
Despite these strengths, the manuscript suffers from a recurring and substantial conceptual issue: systematic over-interpretation of model-data correspondence. While the modeling results are potentially insightful, the extent to which they are presented as recapitulating real claustral neural mechanisms goes beyond what the available data can support.
A fundamental limitation is that the RNN is trained solely on behavioral output, without being constrained by neural data at either single-unit or population levels. As a result, the internal network dynamics are underdetermined and non-unique. Many distinct internal solutions could plausibly generate identical behavior. However, the manuscript frequently treats the specific internal solution discovered in the RNN as if it were a close approximation of the actual claustrum circuit.
This issue is compounded by the sparse nature of the in vivo data used for comparison. The GPFA-based trajectory analyses rely on pseudo-populations and single-trial recordings, yet are interpreted as evidence for robust population-level dynamics. Because neurons were not recorded simultaneously, the inferred trajectories necessarily lack true population covariance and shared trial-to-trial variability, limiting their interpretability as genuine population dynamics. Similarly, conclusions about trajectory-based versus attractor-based computation are drawn almost exclusively from model analyses and then generalized to the biological system.
Overall, while the modeling framework is appropriate as a hypothesis-generating tool, the manuscript repeatedly crosses the line from proposing plausible mechanisms to asserting explanatory or even causal equivalence between the model and the brain. This undermines the otherwise strong contributions of the work.
Below are several specific points that warrant further clarification or revision:
(1) Tone of model-data correspondence
Numerous statements describe the RNN as "closely mimicking," "recapitulating," or being "nearly identical" to claustral neural dynamics, sometimes extending to claims about causal relationships between neural activity and behavior. Given that neural data were not used to train the model, and that only a small subset of trained networks showed the reported dynamics, these statements should be substantially softened throughout the manuscript. The RNN should be framed as providing one possible computational realization consistent with existing data, not as a close instantiation of the biological circuit
(2) Non-uniqueness of RNN solutions
The fact that only a small fraction of trained networks exhibited "claustrum-like" clusters deserves deeper discussion. This observation raises the possibility that the identified solution is fragile or highly specific rather than canonical. The authors should explicitly discuss the non-uniqueness of internal solutions in behavior-trained RNNs, including the range of alternative network dynamics that can reproduce the same behavior. In particular, it should be clarified why the specific network exhibiting "claustrum-like" clusters is informative about claustral computation, rather than representing one arbitrary solution among many.
(3) GPFA trajectory comparisons
The qualitative similarity between RNN trajectories and GPFA-derived trajectories from sparse in vivo data is interesting but insufficient to support claims of robustness or population-level structure. Statements suggesting that these patterns are unlikely to arise from noise or random fluctuations are not justified, given the single-trial, pseudo-population nature of the data. Either additional quantitative controls should be added, or the interpretation should be substantially tempered.
(4) Scope of functional claims
The discussion connecting the findings to broad theories of claustral function, global workspace, or consciousness extends well beyond the data presented. These speculative links should be clearly labeled as such and significantly reduced in strength and prominence.
(5) Comment on Conceptual Interpretation of the Behavioral Paradigm:
The manuscript repeatedly describes the delayed escape task as an "inference-based behavioral paradigm" and states that animals "infer that a value-neutral alternative space is likely to be safer" when the CS is presented in a novel environment. While I appreciate that the US-CS association was established in a different context and that the CS is then presented in a new environment, I am not convinced that the current behavioral evidence uniquely supports an inference interpretation.
First, it is not clear that this task is widely recognized in the literature as a canonical inference task, in the sense of, for example, sensory preconditioning, transitive inference, or model-based inference paradigms. Rather, the observed effect-that CS animals escape faster to a neutral compartment than neutral-CS controls-can be parsimoniously interpreted in terms of generalized threat value, heightened fear/anxiety, or a bias toward avoidance/escape under elevated threat, without requiring an explicit inferential step about the specific safety of the alternative compartment. The fact that no prior training is needed is compatible with flexible generalization, but does not by itself demonstrate inference in a more formal computational sense.
Second, the inference claim becomes central to the manuscript's conceptual framing (e.g., the idea that rsCla supports "inference-based escape"), yet the behavioral analyses presented here and in the cited prior work do not clearly rule out simpler accounts. Clarifying this distinction would help avoid overstating both the inferential nature of the behavior and the specific role of rsCla and the RNN's "claustrum-like" cluster in supporting inference per se, as opposed to more general integration of threat-related signals with an opportunity for escape.
Overall Assessment:
This manuscript presents an interesting and potentially valuable modeling-based framework for thinking about temporal integration in the claustrum, supported by solid slice physiology. However, in its current form, it overstates the degree to which the proposed RNN dynamics reflect actual claustral neural mechanisms. With substantial revision - especially a more cautious interpretation of model-data similarity and a clearer articulation of modeling limitations - the study could make a meaningful contribution as a hypothesis-generating work rather than a definitive mechanistic account.
-
Reviewer #2 (Public review):
This manuscript reports the behavior of a computational model of rat claustral neurons during the performance of a behavioral task known as the delayed escape task (in this reviewer's understanding, this behavioral task was created and implemented by this group only). These authors have argued in a prior manuscript (Han et al.) that a group of neurons located "rostral to striatum" is part of the claustrum. The group names the region the "rostral to striatum claustrum." Additionally, in the Han et al. paper, the authors argue that these cells are responsible for maintaining a signal that lasts through the delay period.
The main findings of the current paper are:
(1) The authors have built a model network that was trained to show firing similar to what was reported for rats in their prior paper.
(2) The …
Reviewer #2 (Public review):
This manuscript reports the behavior of a computational model of rat claustral neurons during the performance of a behavioral task known as the delayed escape task (in this reviewer's understanding, this behavioral task was created and implemented by this group only). These authors have argued in a prior manuscript (Han et al.) that a group of neurons located "rostral to striatum" is part of the claustrum. The group names the region the "rostral to striatum claustrum." Additionally, in the Han et al. paper, the authors argue that these cells are responsible for maintaining a signal that lasts through the delay period.
The main findings of the current paper are:
(1) The authors have built a model network that was trained to show firing similar to what was reported for rats in their prior paper.
(2) The authors' analysis of model behavior is used to suggest that the model network recapitulates biological activity, including the existence of a cluster of cells mainly responsible for the delay period firing.
(3) The authors offer evidence from patch clamp recordings for excitatory interconnections among claustral neurons that are an essential feature of the model network.
A major value of the computational network is that "trials" of the network can be performed. In experiments on animals, only single trials can be used.
Concerns:
(1) This paper is based on behavioral results and neural recordings from their prior paper (Han et al.), but data, e.g., in Figure 1, are not clearly identified as new or as coming from that source. Figure 1A, for example, appears to be taken directly from Han et al. No methods are given in this manuscript for the behavioral testing or the in vivo electrophysiology.
(2) Many other details are unclear. Examples include model training, the weight matrices and how these changed with training (p. 13), equations 2 and 3 (p. 13), the sources for the constants in the equations (p. 14), the methods (anesthesia, stereotaxic coordinates, injection specifics and details for "sparse expression") for the ChrimsonR injections.
(3) The explorations of model behavior are a catalog of everything tried rather than an organized demonstration of what the model can and cannot do. The figures could be reduced in number to emphasize the key comparisons of the different clusters and the model's behavior under different conditions, intended to "test" the model.
(4) On page 6, the E-E connectivity is argued from Shelton et al. (2025) and against Kim et al. (2016), but ignores Orman (2015), which, to this reviewer's knowledge, was the first to demonstrate such connectivity, including the long-duration events and impact of planes of section.
(5) Whereas the authors are entitled to their own opinion of prior work (references 3-8), it is inappropriate to misrepresent prior work as only demonstrating a "limited function" of claustum. Additional papers by Mathur's group and Citri's group are ignored.
In summary, the authors have made a computational model that recapitulates the firing of a subset of potentially claustral neurons during a particular behavioral task (delayed escape is certainly not the only behavior that involves claustrum - see e.g., attention, salience, sleep). If the conclusion is that excitatory claustral cells must be connected to other excitatory claustral cells, such a conclusion is not new, and the electrophysiological E-E metrics are not well quantified (e.g., connectivity frequency, strength of connection). If the model is intended to predict how the claustrum might accomplish any other task, there is insufficient detail to evaluate the model beyond the evidence that the model creates a subset of cells that can sustain firing during the delay period in the delayed escape task.
All relevant work must be appropriately cited throughout the manuscript.
-
Author response:
Public Reviews:
Reviewer #1 (Public review):
Our goal was to propose a possible computational mechanism underlying information integration in the claustrum, not to claim structural or causal equivalence between the model and the biological circuit. We acknowledge that some expressions in the original manuscript may have been interpreted as exceeding this intention, and we will revise the text to explicitly soften such statements.
It is well established that behavior-trained RNNs can admit multiple internal solutions capable of producing the same behavioral output, and we fully agree with this point. Among the many possible solutions, we focused on networks that exhibited dynamical properties consistent with independently obtained behavioral and physiological findings. Thus, in our view, biological plausibility in this …
Author response:
Public Reviews:
Reviewer #1 (Public review):
Our goal was to propose a possible computational mechanism underlying information integration in the claustrum, not to claim structural or causal equivalence between the model and the biological circuit. We acknowledge that some expressions in the original manuscript may have been interpreted as exceeding this intention, and we will revise the text to explicitly soften such statements.
It is well established that behavior-trained RNNs can admit multiple internal solutions capable of producing the same behavioral output, and we fully agree with this point. Among the many possible solutions, we focused on networks that exhibited dynamical properties consistent with independently obtained behavioral and physiological findings. Thus, in our view, biological plausibility in this study is not grounded in structural isomorphism, but rather in whether the core population-level dynamical properties observed in the model are reproducible in actual claustral population activity.
We also agree with the reviewer that our original qualitative comparison of GPFA-based latent trajectories did not provide sufficient quantitative support. In the revised manuscript, we have therefore added an eigenvalue-based quantitative analysis of the dimensional structure of population trajectories. This analysis does not depend on the identity of the dimensionality-reduction method itself, but instead focuses on quantifying the geometric structure of population-state trajectories as they evolve over time. Applying the same metric to both the RNN and biological claustrum data revealed consistent condition-specific differences in population dynamics.
This quantitative addition strengthens the previous qualitative trajectory comparison and clarifies that the model implements a specific computational dynamical regime that directionally corresponds to claustral population activity. While this does not imply uniqueness of the model, we believe it suggests that the proposed computational principle represents a biologically realizable candidate mechanism.
(1) Tone of model-data correspondence
Numerous statements describe the RNN as "closely mimicking," "recapitulating," or being "nearly identical" to claustral neural dynamics, sometimes extending to claims about causal relationships between neural activity and behavior. Given that neural data were not used to train the model, and that only a small subset of trained networks showed the reported dynamics, these statements should be substantially softened throughout the manuscript. The RNN should be framed as providing one possible computational realization consistent with existing data, not as a close instantiation of the biological circuit.
We agree with the reviewer’s concern. Expressions such as “closely mimicked,” “nearly identical,” and “recapitulate” will be replaced with more moderate language.
(2) Non-uniqueness of RNN solutions
The fact that only a small fraction of trained networks exhibited "claustrum-like" clusters deserves deeper discussion. This observation raises the possibility that the identified solution is fragile or highly specific rather than canonical. The authors should explicitly discuss the non-uniqueness of internal solutions in behavior-trained RNNs, including the range of alternative network dynamics that can reproduce the same behavior. In particular, it should be clarified why the specific network exhibiting "claustrum-like" clusters is informative about claustral computation, rather than representing one arbitrary solution among many.
As the reviewer noted, behavior-trained RNNs can yield multiple internal solutions that generate the same behavioral output, and we acknowledge this non-uniqueness. However, we do not interpret the relatively low success rate (5/100 networks) as evidence of fragility. Rather, we interpret it as suggesting that the emergence of this particular dynamical regime requires stringent structural constraints.
The computational demands of the task—specifically, the integration of temporally separated signals—drive convergence toward networks capable of sustaining persistent activity through recurrent excitatory connectivity. Indeed, all networks exhibiting a claustrum-like cluster shared a strong recurrent excitatory structure within Cluster 1, a structural feature consistent with our slice electrophysiology findings.
Our criterion for selecting RNNs was their ability to reproduce behavioral and physiological observations from the delayed escape experiment. Excluded RNNs may reflect alternative information-processing strategies characteristic of other brain regions or artificial logical solutions. Importantly, claustrum-like dynamics were not explicitly enforced during training; they emerged spontaneously under behavioral constraints, suggesting that this solution is not arbitrary.
Furthermore, the computational principles derived from the RNN were quantitatively consistent with in vivo single-neuron activity. Using an eigenvalue-based metric (λ3/Σλ), both the RNN and biological claustrum data showed effects in the same direction. Leave-one-neuron-out analyses further demonstrated that this pattern was broadly distributed across neurons in the claustrum. These convergent results suggest that the identified network captures a computational regime that is consistent with claustral population dynamics, rather than representing an arbitrary solution unrelated to the biological observations.
(3) GPFA trajectory comparisons
The qualitative similarity between RNN trajectories and GPFA-derived trajectories from sparse in vivo data is interesting but insufficient to support claims of robustness or population-level structure. Statements suggesting that these patterns are unlikely to arise from noise or random fluctuations are not justified, given the single-trial, pseudo-population nature of the data. Either additional quantitative controls should be added, or the interpretation should be substantially tempered.
We agree that the original GPFA trajectory comparison in the biological claustrum data remained qualitative and did not sufficiently establish robustness or population-level structure. We have therefore added quantitative analyses in the revised manuscript.
Before presenting these analyses, we clarify methodological limitations inherent in pseudopopulation and single-trial data. GPFA estimates latent trajectories based on covariance structure and temporal smoothness assumptions. In pseudopopulations, true simultaneously recorded covariance cannot be fully reconstructed. Although our dataset is based on single trials rather than trial-to-trial variability, we acknowledge that latent-space estimation depends on covariance structure.
Therefore, the additional quantitative metric is not independent of the GPFA estimation stage; rather, it evaluates the geometric structure of single-trial latent trajectories estimated by GPFA.
Specifically, for biological data, we reanalyzed GPFA-estimated latent trajectories in PCA space and computed an eigenvalue-based metric (λ3/Σλ). Across 20 time bins, a sliding window of 10 bins was applied. For each window, we computed the covariance matrix and extracted eigenvalues for PC1, PC2, and PC3. The third eigenvalue (λ3) was normalized by total variance (Σλ = λ1 + λ2 + λ3). This metric quantifies the extent to which trajectories deviate from a planar (two-dimensional) structure into a third dimension. An increase in λ3/Σλ indicates the formation of a higher-dimensional geometric structure.
For RNN data, since all unit activities were simultaneously observed and sufficient trials were available, we directly applied PCA to population activity without GPFA. Mean trajectories across trials were computed, and the same λ3/Σλ metric was applied. Although the initial dimensionality-reduction steps differ, the final metric definition and computation are identical. Thus, the comparison focuses on geometric dimensional structure rather than the dimensionality-reduction method itself.
Importantly, within the biological dataset, GPFA estimation, preprocessing, pseudopopulation construction, subsampling strategy, temporal alignment, and smoothing were applied identically across the CS and Neutral conditions. Under this common analysis framework, λ3/Σλ values were consistently higher in the CS condition than in the Neutral condition.
For the RNN data, an identical analysis pipeline was applied across the CS+Open and Open-only conditions. In this case as well, λ3/Σλ values were significantly higher in the CS+Open condition than in the Open-only condition.
If structural bias arose from covariance estimation or dimensionality reduction, it would be expected to affect conditions similarly within each dataset. The observation that λ3/Σλ increases selectively in the CS condition in biological data and in the CS+Open condition in the RNN therefore supports the interpretation that the effect reflects a condition-specific dynamical difference rather than an artifact of dimensionality reduction.
To further examine whether the effect was driven by a small subset of neurons, we performed leave-one-neuron-out analyses in the biological dataset. In the CS group, most neurons contributed relatively evenly to the metric, whereas such distributed contribution was not observed in the Neutral group. This suggests that the three-dimensional structure reflects an organized population-level phenomenon rather than covariance dominated by a small number of outlier neurons.
These results indicate that the consistent elevation of λ3/Σλ in the CS condition (biological data) and in the CS+Open condition (RNN) reflects a genuine dynamical feature rather than an artifact arising from pseudopopulation construction or dimensionality reduction.
Taken together, the three-dimensional geometric structure observed in GPFA-based latent trajectories is unlikely to reflect random noise. The replication of the same quantitative metric in the RNN, using an independent dimensionality-reduction procedure, strengthens the correspondence between the two systems. We appreciate the reviewer’s suggestion for quantitative reinforcement, which has substantially strengthened the manuscript.
(4) Scope of functional claims
The discussion connecting the findings to broad theories of claustral function, global workspace, or consciousness extends well beyond the data presented. These speculative links should be clearly labeled as such and significantly reduced in strength and prominence.
We agree with the reviewer and will clearly indicate that references to broader theoretical interpretations are speculative. We will substantially reduce their strength and emphasis.
(5) Comment on Conceptual Interpretation of the Behavioral Paradigm:
The manuscript repeatedly describes the delayed escape task as an "inference-based behavioral paradigm" and states that animals "infer that a value-neutral alternative space is likely to be safer" when the CS is presented in a novel environment. While I appreciate that the US-CS association was established in a different context and that the CS is then presented in a new environment, I am not convinced that the current behavioral evidence uniquely supports an inference interpretation.
We agree with the reviewer’s concern. We will describe the delayed escape task as “a behavioral paradigm that requires integration of temporally separated task-relevant signals” and remove inference-related terminology throughout the manuscript.
Reviewer #2 (Public review):
We appreciate the reviewer’s constructive and well-balanced comments. We regret that some of our wording and the scope of our introduction and discussion may not have appropriately reflected the contributions of prior studies. We will revise the manuscript accordingly to ensure that previous literature is more accurately and fairly acknowledged. In addition, we will reorganize the figures to more clearly present the hypotheses being tested and will provide additional details regarding both the modeling framework and the experimental procedures.
(1) This paper is based on behavioral results and neural recordings from their prior paper (Han et al.), but data, e.g., in Figure 1, are not clearly identified as new or as coming from that source. Figure 1A, for example, appears to be taken directly from Han et al. No methods are given in this manuscript for the behavioral testing or the in vivo electrophysiology.
We will clarify more explicitly which data and methods originate from Han et al. (2024). In the original manuscript, Figure 1 panels A, D, E, F, and L (left) were indicated in the legend as originating from Han et al. (2024). We will further clarify this distinction in the main text. Additionally, we will briefly describe the behavioral experiments and in vivo electrophysiology performed in Han et al. in the Methods section, with appropriate citation.
(2) Many other details are unclear. Examples include model training, the weight matrices and how these changed with training (p. 13), equations 2 and 3 (p. 13), the sources for the constants in the equations (p. 14), the methods (anesthesia, stereotaxic coordinates, injection specifics and details for "sparse expression") for the ChrimsonR injections.
As requested, we will provide additional details regarding model training procedures, weight matrices and their evolution during training, equations (2) and (3), the origin of constants used in the equations, and detailed methods for ChrimsonR injection (anesthesia, stereotaxic coordinates, injection parameters, and clarification of “sparse expression”).
(3) The explorations of model behavior are a catalog of everything tried rather than an organized demonstration of what the model can and cannot do. The figures could be reduced in number to emphasize the key comparisons of the different clusters and the model's behavior under different conditions, intended to "test" the model.
We will reorganize the figures to emphasize core results and clarify that the primary goal is to test and validate the computational model.
(4) On page 6, the E-E connectivity is argued from Shelton et al. (2025) and against Kim et al. (2016), but ignores Orman (2015), which, to this reviewer's knowledge, was the first to demonstrate such connectivity, including the long-duration events and impact of planes of section.
We will cite Orman (2015) as suggested and note that persistent activity has been observed in slices cut at specific angles, consistent with our findings.
(5) Whereas the authors are entitled to their own opinion of prior work (references 3-8), it is inappropriate to misrepresent prior work as only demonstrating a "limited function" of claustum. Additional papers by Mathur's group and Citri's group are ignored.
We will remove wording implying “limited” prior work and appropriately acknowledge contributions from the Mathur and Citri groups.
In summary, the authors have made a computational model that recapitulates the firing of a subset of potentially claustral neurons during a particular behavioral task (delayed escape is certainly not the only behavior that involves claustrum - see e.g., attention, salience, sleep). If the conclusion is that excitatory claustral cells must be connected to other excitatory claustral cells, such a conclusion is not new, and the electrophysiological E-E metrics are not well quantified (e.g., connectivity frequency, strength of connection). If the model is intended to predict how the claustrum might accomplish any other task, there is insufficient detail to evaluate the model beyond the evidence that the model creates a subset of cells that can sustain firing during the delay period in the delayed escape task.
Across all whole-cell recordings, optogenetic responses were observed in 38 out of 43 patched cells (~90%), suggesting that a high proportion of claustral neurons receive intra-claustral excitatory input. However, precise connectivity frequency and strength cannot be determined from the current dataset.
As the reviewer noted, our RNN is specialized for the delayed escape task, and we do not claim direct generalization to other proposed claustral functions such as attention, salience, or sleep. The goal of this study is to computationally characterize the temporal integration mechanism observed in this specific task.
While our model is specific to the delayed escape task, the computational principle identified here—nonlinear trajectory-based temporal integration supported by recurrent excitatory connectivity—may represent a more general mechanism for integrating temporally separated signals. However, testing such generality lies beyond the scope of the present study and will be framed as a future direction in the revised Discussion.
-
