Sexual dimorphism in obesity is governed by RELMα regulation of adipose macrophages and eosinophils
Curation statements for this article:-
Curated by eLife

eLife assessment
In this study, Li and al describe valuable insights into the mechanisms underlying sex-differences diet-induced obesity in mice, with a role of macrophage-derived RELMa secretion in female-specific protection. They provide solid evidence for the impact of RELMa signaling in eosinophil recruitment for diet-induced obesity protection in female mice. Single-cell RNA-seq analysis of the stromal vascular fraction of control and RELMa deficient animals methods were used to investigate molecular mechanisms underlying the protection as a powerful method, although the analysis of this data is difficult to evaluate with incomplete methodological information.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
- Preprint highlights for July (scietyHQ)
Abstract
Obesity incidence is increasing worldwide with the urgent need to identify new therapeutics. Sex differences in immune cell activation drive obesity-mediated pathologies where males are more susceptible to obesity comorbidities and exacerbated inflammation. Here, we demonstrate that the macrophage-secreted protein RELMα critically protects females against high-fat diet (HFD)-induced obesity. Compared to male mice, serum RELMα levels were higher in both control and HFD-fed females and correlated with frequency of adipose macrophages and eosinophils. RELMα-deficient females gained more weight and had proinflammatory macrophage accumulation and eosinophil loss in the adipose stromal vascular fraction (SVF), while RELMα treatment or eosinophil transfer rescued this phenotype. Single-cell RNA-sequencing of the adipose SVF was performed and identified sex and RELMα-dependent changes. Genes involved in oxygen sensing and iron homeostasis, including hemoglobin and lncRNA Gm47283/Gm21887, correlated with increased obesity, while eosinophil chemotaxis and response to amyloid-beta were protective. Monocyte-to-macrophage transition was also dysregulated in RELMα-deficient animals. Collectively, these studies implicate a RELMα–macrophage–eosinophil axis in sex-specific protection against obesity and uncover new therapeutic targets for obesity.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
In this manuscript, Li et al characterize sex differences in the impact of macrophage RELMa in protection against diet-induced obesity [DIO]. This is a key area of interest as obesity studies in mice have generally focused exclusively on male animals, as they tend to gain more weight, faster than female mice. The authors use a combination of flow cytometry, adoptive transfer, and single-cell transcriptomics to characterize the mechanism of action for female-specific DIO protection. They identify a potential role for eosinophils in mediating female DIO protection downstream of RELMa production by macrophage. They also use the transcriptomic characterization of the stromal vascular fraction of the adipose tissue to evaluate molecular and cellular drivers of this sex-specific DIO protection.
Author Response
Reviewer #1 (Public Review):
In this manuscript, Li et al characterize sex differences in the impact of macrophage RELMa in protection against diet-induced obesity [DIO]. This is a key area of interest as obesity studies in mice have generally focused exclusively on male animals, as they tend to gain more weight, faster than female mice. The authors use a combination of flow cytometry, adoptive transfer, and single-cell transcriptomics to characterize the mechanism of action for female-specific DIO protection. They identify a potential role for eosinophils in mediating female DIO protection downstream of RELMa production by macrophage. They also use the transcriptomic characterization of the stromal vascular fraction of the adipose tissue to evaluate molecular and cellular drivers of this sex-specific DIO protection.
Although the authors provide solid evidence for many claims in the manuscript, there is generally not enough information about the studies' methods (especially on the computational/data analysis aspects) for a careful evaluation of the result's robustness at this stage.
We have significantly expanded the methodology, especially of the scRNAseq, and deposited the script and raw data in public repositories. We also validated our methods and can confirm that the analysis presented is robust. This resubmission contains new Fig 7 and new supplementary material with this methodology and validation.
Reviewer #2 (Public Review):
In the study by Li et al., the authors hypothesize that RELMa, a macrophage-derived protein, plays a sex-dimorphic role as a protective factor in obesity in females vs males. The authors perform largely in vivo studies utilizing male and female WT and RELMa KO mice on a high-fat diet and perform an in-depth analysis of immune cell composition, gene expression, and single-cell RNA Sequencing. The authors find that WT females are protected from obesity and inflammation vs males, and this protection is lost in female RELMa KO mice. Further analysis by the authors including flow cytometry of the visceral fat SVF in female WT mice showed reduced macrophage infiltration, higher levels of eosinophils, and Th2 cytokine expression compared to WT male mice and female KO mice. The authors show that protection from obesity and inflammation in female RELMa KO mice can be rescued with an injection of eosinophils and recombinant RELMa. Lastly, the authors use single-cell RNA-Sequencing to further analyze SVF cells in WT and KO male and female mice on a high-fat diet.
Overall, we find that the study represents an important finding in the immunometabolism field showing that RELMa is a key myeloid-derived factor that helps influence the macrophage-eosinophil function in female mice and protects from diet-induced obesity and inflammation in a sexually dimorphic manner. Overall, the study provides strong and convincing data supporting the authors' hypothesis and conclusion.
We thank the reviewer for their positive review of our manuscript and their helpful feedback which we address below.
Reviewer #3 (Public Review):
Li, Ruggiero-Ruff et al. examine the role of RELMα, an anti-inflammatory macrophage signature gene, in mediating sex differences in high-fat diet (HFD)-induced obesity in young mice. Specifically, the authors hypothesize that RELMα protects females against HFD-induced obesity. Comparisons between RELMα-knockout (KO) and wildtype (WT) mice of both sexes revealed sex- and RELMα-specific differences in weight gain, immune cell populations, and inflammatory signaling in response to HFD. RELMα-deficiency in females led to increased weight gain, expansion of pro-inflammatory macrophage populations, and eosinophil loss in response to HFD. Female RELMα-deficiency could be rescued by RELMα treatment or eosinophil transfer. Single-cell RNA-sequencing (scRNA-seq) of adipose stromal vascular fraction (SVF) revealed sex- and RELMα-dependent differences under HFD conditions and identified potential "pro-obesity" and "anti-obesity" genes in a cell-type-specific manner. Using trajectory analysis, the authors suggest dysregulation of macrophage-to-monocyte transition in RELMα-deficient mice.
The conclusions of this paper are mostly well supported by the data, but some aspects of the statistical and single-cell analyses will need to be corrected, clarified, and extended to enhance the report.
We thank Dr. Ocanas for their positive comments and for the helpful feedback to improve our study. We have addressed all the comments and significantly revised the manuscript.
Strengths:
The authors use several orthogonal approaches (i.e., flow cytometry, immunohistochemistry, scRNA-Seq) and models to support their hypotheses.
The authors demonstrate that phenotypes observed in HFD-fed females with RELMα-deficiency (i.e., weight gain, loss of eosinophils, a gain of M1 macrophages) can be rescued by RELMα treatment or eosinophil transfer.
The authors recognized the complexity of macrophage activation that is beyond the 'M1/M2' paradigm and informed readers in the introduction as to why this paradigm was used in this study. During the scRNA-seq analyses, the authors further sub-cluster macrophages to include more granularity.
Weaknesses:
- There are several instances in the text where the authors claim that there is a significant difference between the two groups, but the statistics for these comparisons are not shown in the figure.
Because we are dealing with three variables: genotype, diet and sex, and many differences, we thought it too complicated to add all the significant differences on the graph, but sometimes just mentioned these in the text with a p value, or didn’t mention at all if the difference was obvious, or not meaningful (for example, we weren’t interested in comparing a WT male on a Ctr diet with a RELMalpha KO female on a HFD for the purpose of our hypothesis). We have now ensured clarity in the text and in the figures, and addressed the specific point-by-point comments from the reviewer. We have also now carefully re-evaluated the text to ensure that any significant differences we discuss are shown in the figure.
- It is unfortunate that eosinophils could not be identified in the single-cell analysis since this population of cells was shown to be important in rescuing the RELMα-deficiency in HFD-fed females. The authors should note in the discussion how future scRNA-Seq experiments could overcome this limitation (i.e., enriching immune cells prior to scRNA-Seq).
We were indeed disappointed that we were not able to obtain eosinophil single cell seq, but realize that this is a reported issue in the field. We have expanded our discussion of this and cited a paper that performs eosinophil single cell sequencing (published at the time our manuscript was being submitted): ““At the same time as our ongoing analysis, the first publication of eosinophil single cell RNA-seq was published, using a flow cytometry based approach rather than 10x, including RNAse inhibitor in the sorting buffer, and performing prior eosinophil enrichment (PMID: 36509106). Based on guidance from 10x, we employed targeted approaches to identify eosinophil clusters according to eosinophil markers (e.g. Siglecf, Prg2, Ccr3, Il5r), and relaxed the scRNA-Seq cutoff analysis to include more cells and intronic content, but still could not find eosinophils. We conclude that eosinophils may be absent due to the enzyme digestion required for SVF isolation and processing for single cell sequencing, which could lead to specific eosinophil population loss due to low RNA content, RNases or cell viability issues. Future experiments would be needed to optimize eosinophil single cell sequencing, based on the recent publication of eosinophil single cell sequencing.”
3a) There are several issues with the scRNA-Seq analysis and interpretation. More details on the steps taken in the single-cell analyses should be included in the methods section.
We agree with the reviewer that more details on steps taken in the single cell data processing and bioinformatics needs to be included in the methods section. We included more information and separated sections within the data processing section in the Materials and Methods on the methodology used for these approaches, as well as provided a code for our data processing in a public Github repository: https://github.com/rrugg002/Sexual-dimorphism-in-obesity-is-governed-by-RELM-regulation-of-adipose-macrophages-and-eosinophils.
b) With regards to the 'pseudobulk' analyses presented in Figs. 5-6, several of the differentially expressed genes identified in Fig. 6 are hemoglobin genes (i.e., Hba, Hbb genes). It is not uncommon to filter these genes out of single-cell analysis since their presence usually indicates red blood cell (RBC) contamination (PMID: 31942070, PMID: 35672358). We would recommend assessing RBC contamination as well as removing Fig. 6 from the manuscript and focusing on cell-type-specific analyses. Re-analysis will likely have an impact on the overall conclusions of the study.
Prior to our first submission, we consulted with 10x support scientists and the UCR bioinformatics core director to ensure that our analysis included the appropriate filtering. We have now added details in the Methods. The PMIDs provided above are from studies that looked at hippocampus development (where they didn’t perfuse so there may be blood contamination) or whole blood (where there would be significant red blood cell contamination). In contrast, we perfused our mice and treated the single cell suspension with RBC lysis buffer, as detailed in Methods. Also, we have now extended our scSeq analysis to compare hemoglobin RNA to red blood cell specific markers including Gypa/CD235a. While hemoglobin is distributed throughout the myeloid population in the female KO mice, Gypa/CD235a, which would suggest RBC contamination is not expressed at all (see new Fig 7B). Additionally, we provide hemoglobin protein ELISA and IF staining to support our finding that macrophages from KO mice express hemoglobin protein. Last, two publications support hemoglobin expression by nonerythroid sources, including macrophages (PMID: 10359765; PMID: 25431740). While we are confident based on above that our data is not due to RBC contamination, we cannot exclude the fact that, although unlikely, macrophages may be phagocytosing RBC and preserving specifically hemoglobin RNA and protein. Nonetheless, we discuss this possibility in the text. In conclusion, based on the justification above and the new data, we are confident that our findings and overall conclusions are robust.
To assess for potential RBC contamination, in addition to Gypa, we additionally looked at top genes expressed by murine erythrocytes (PMID: 24637361). Please see below feature plots, showing little to no expression, and a very different distribution than the hemoglobin genes (see new Fig 7a):
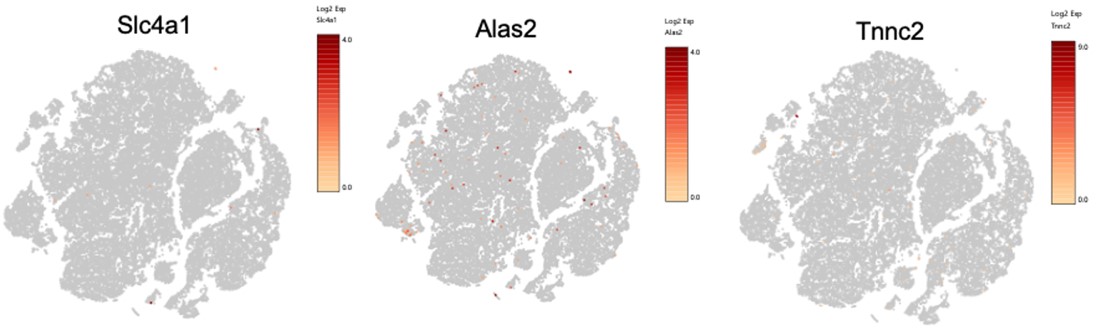 Also, we had a small cluster of potential RBCs (only 75 cells) that we filtered out of downstream DEG analysis, which revealed the same data as in the first submission.
Also, we had a small cluster of potential RBCs (only 75 cells) that we filtered out of downstream DEG analysis, which revealed the same data as in the first submission.- Within the text, there are several instances where the authors claim that a pathway is upregulated based on their Gene Ontology (GO) over-representation analysis (ORA). To come to this conclusion, the authors identify genes that are upregulated in one condition and then perform GO-ORA on these genes. However, the authors do not consider negative regulators, whose upregulation would actually decrease the pathway. Authors should either replace their GO-ORA analysis with one that considers the magnitude and direction of differentially expressed genes and provides an activation z-score (i.e., Ingenuity Pathway Analysis) or replace instances of 'upregulated' or 'downregulated' pathways with 'over-represented' pathways.
Unfortunately, we did not have access to IPA for this project, therefore we have changed our analysis to over and under-represented pathways as suggested.
- For Fig.7A, a representative tSNE plot for each group (WT Female, KO Female, WT Male, KO Male) should be shown to ensure there is proper integration of the clusters across groups. There are some instances where the scRNA-Seq data do not appear to be integrated properly (i.e., Supplemental Figure 2C). The authors should explore integration techniques (i.e., Seurat; PMID: 29608179) to correct for potential batch effects within the analysis.
We thank the reviewer for the suggestion of proper integration of the clusters across groups. We performed integration using the Cell Ranger aggregation (aggr) pipeline (see updated materials and methods section). In addition, many technical controls were performed to prevent batch effects between our samples. For sequencing, we used the 10x genomics library sequencing depth and run parameters for both gene expression and multiplexing libraries. For all 3’ gene expression library sequencing, we sequenced at a depth of 20,000 read pairs per cell and for all cell multiplexing library sequencing we sequenced at a depth of 5,000 read pairs per cell. All libraries were paired-end dual indexed libraries and were pooled on one flow cell lane using a 4:1 ratio (3’ Gene expression: Multiplexing ratio) in the Novaseq, as recommended by 10x Genomics, in order to maintain nucleotide diversity and prevent batch effects during the sequencing process. When performing integration/aggregation of all sample gene expression libraries using the Cell Ranger aggregation (aggr) pipeline, we performed sequencing depth normalization between all samples. Cell Ranger does this by equalizing the average read depth per cell between groups before merging all sample libraries and counts together. This is a default setting in the Cell Ranger aggr pipeline, and this approach avoids artifacts that may be introduced due to differences in sequencing depth. Thus, we are confident that changes we observed in gene expression and cell type populations are due to biological differences and not technical variability. Below we have provided a tSNE plot showing clustering of all 12 samples after we performed integration:
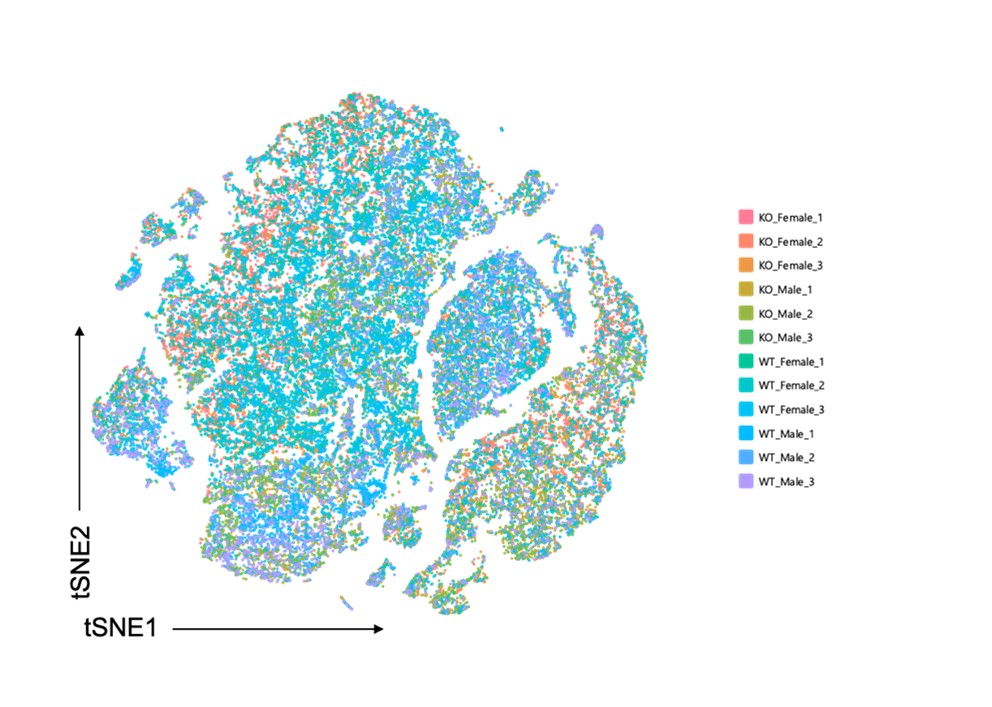
We updated old Fig.7 (now Fig. 6) and included a representative tSNE plot for each group. We also updated the tSNE plot for Figure 5-figure supplement 2C (previously S2C) showing overall clustering amongst all groups. The largest population differences occurred in the fibroblast population and these population differences were largely due to sex differences. Because we are confident that integration was performed appropriately and that batch effects were controlled for, we believe these sex differences are a biological effect.
- LncRNA Gm47283 is identified as a gene that is differentially expressed by genotype in HFD females (Fig. 7G); however, according to Ensembl this gene is encoded on the Y-chromosome (https://uswest.ensembl.org/Mus_musculus/Gene/Summary?g=ENSMUSG00000096768;r=Y:90796007-90827734). The authors should use the RELMα genotype and sex chromosomally-encoded genes to confirm that their multiplexing was appropriate.
We agree with the reviewer that it is crucial to confirm that multiplexing and all subsequent analyses are performed correctly. Comparison between males and females contains internal controls that increase confidence, such as Xist gene that is expressed only in females, and Ddx3y that is located on the Y chromosome. LncRNA, Gm47283 is located in the syntenic region of Y chromosome and is also present in females, annotated as Gm21887 located in the syntenic region of the X chromosome. It also has 100% alignment with Gm55594 on X chromosome. Additionally, it is also referred to erythroid differentiation regulator 1 (Erd1), x or y depending on the chromosome, although NCBI database specifies partial assembly and incomplete annotation. Therefore, this explains why we see expression of this gene in females. We have discussed this in the text. We revised the text to refer to this LncRNA as Gm47283/Gm21887 to prevent further confusion. The RELMalpha genotype (absence in the KO) was also confirmed. Last, the PC analysis (see Fig 5) supports clustering by group.
- For Fig. 8, samples should be co-clustered and integrated across groups before performing trajectory analysis to allow for direct comparisons between groups.
We appreciate the valuable feedback and suggestions, which have been helpful in clarifying the trajectory analysis, which we have done as follows:
Regarding the co-clustering and integration of our samples across groups, here is the explanation of our trajectory analysis approach. We have co-clustered all of our samples using the align_cds function from the Monocle3 package. We have included the code for Figure 8 in our Github repository at https://github.com/rrugg002/Sexual-dimorphism-in-obesity-is-governed-by-RELM-regulation-of-adipose-macrophages-and-eosinophils/blob/main/Figure8.R. Specifically, lines 138, 166, 196 and 225 of the code indicate that the align_cds function was used to cluster our samples by "Sample.ID".
The align_cds function in Monocle3 can be used to co-cluster all samples in a single-cell RNA-seq experiment by aligning coding sequences (CDS) across different cell types or conditions. The align_cds function takes a set of reference CDS sequences and single-cell RNA-seq reads and identifies the CDS sequences within each read, allowing the identification of differentially expressed genes across different cell types or conditions based on the aligned CDS sequences. More details about align_cds can be found here https://rdrr.io/github/cole-trapnell-lab/monocle3/man/align_cds.html .
We hope that this additional information alleviates the reviewer’s concerns.
- Since the experiments presented in this report were from young mice using a single diet intervention, the authors should comment on how age and other obesogenic diets may impact the results found here. Also, the authors should expand their discussion as to what upstream regulators (i.e., hormones or genetics) may be driving the sex differences in RELMα expression in response to HFD.
We thank the reviewer for the suggestion. We included several sentences to address this comment. However, since reviewers commented that some of the text needs to be trimmed down, extensive discussion regarding reasons for sex differences, which are numerous, are outside the scope of this manuscript. For example, sex differences can arise from all or any of these:
Sex steroid hormones (estrogen and testosterone) are an obvious possibility for sex differences and this discussion has been included below and in the text.
Sex differences we observe may stem from variety of other factors, besides ovarian estrogen; including extraovarian estrogen, primarily estrogen produced in adipose tissues (32119876).
Sex differences exist in fat deposition, which may or may not be estrogen dependent (25578600, 21834845).
Sex difference were determined in metabolic rate and oxidative phosphorylation, which may also be independent of estrogen (28650095, and reviewed in 26339468).
Sex differences exist in the immune system, some of which are estrogen independent, but dependent on sex chromosomes (32193609).
Sex differences particularly in myeloid lineage, which may also be estrogen independent (25869128).
Sex differences were determined in adipokine levels, including leptin and adiponectin, which influence immune cells in adipose tissues (33268480).
The role of estrogen is not clear either, and thus extensive discussion is not possible. Numerous studies demonstrated that estrogen is protective from inflammation, thus it is possible that estrogen drives some of the sex differences observed herein. However, several studies determined that estrogen can be pro-inflammatory (20554954, 15879140, 18523261). Previous publications by us (30254630, 33268480) and others (25869128) demonstrated intrinsic sex differences in immune system, that are maybe dependent on sex chromosome complement and/or Xist expression (34103397, 30671059).
Studies are more consistent that estrogen is protective from weight gain: postmenopausal women with diminished estrogen, and ovariectomized animal models gain weight. The effects of ovariectomy on weight gain and its additive effects with high fat diet were reported in Rhesus monkeys (for example PMID: 2663699; and PMID: 16421340); and in rodents (PMID: 7349433).
The reviewer is correct that the effects of aging or estrogen on RELMa levels would be of significant interest, and could be a future direction of our studies. Aging-mediated increase in inflammation (including of adipose tissue, recently reviewed in 36875140), that may be dependent on estrogen, can exacerbate obesity-mediated inflammation. We have added this discussion.
For these reasons we limited our discussion regarding possible differences and stated this in the discussion: “Several studies demonstrated the protective role of estrogen in obesity-mediated inflammation and in weight gain, as discussed above. Whether estrogen protection occurs via estrogen regulation of RELMa levels is a focus of our future studies. Alternatively, intrinsic sex differences in immune system have been demonstrated as well (30254630, 33268480, 25869128) that are dependent on sex chromosome complement and/or Xist expression (34103397, 30671059), and RELMa may be regulated by these as well. Additionally, ageing-mediated increase in inflammation (including of adipose tissue, recently reviewed in 36875140), may also occur via changes in RELMa levels. Our studies used young but developmentally mature mice (4-6 weeks old when placed on diet, 18 weeks old at sacrifice), and future work on aged mice would be needed to investigate aging-mediated inflammation. Furthermore, there are sex differences in fat deposition, metabolic rates and oxidative phosphorylation (reviewed in 26339468), and adipokine expression (Coss) that regulate cytokine and chemokines levels, and therefore may regulate levels of RELMa as well. These possibilities will be addressed in future studies.”
-
eLife assessment
In this study, Li and al describe valuable insights into the mechanisms underlying sex-differences diet-induced obesity in mice, with a role of macrophage-derived RELMa secretion in female-specific protection. They provide solid evidence for the impact of RELMa signaling in eosinophil recruitment for diet-induced obesity protection in female mice. Single-cell RNA-seq analysis of the stromal vascular fraction of control and RELMa deficient animals methods were used to investigate molecular mechanisms underlying the protection as a powerful method, although the analysis of this data is difficult to evaluate with incomplete methodological information.
-
Reviewer #1 (Public Review):
In this manuscript, Li et al characterize sex differences in the impact of macrophage RELMa in protection against diet-induced obesity [DIO]. This is a key area of interest as obesity studies in mice have generally focused exclusively on male animals, as they tend to gain more weight, faster than female mice. The authors use a combination of flow cytometry, adoptive transfer, and single-cell transcriptomics to characterize the mechanism of action for female-specific DIO protection. They identify a potential role for eosinophils in mediating female DIO protection downstream of RELMa production by macrophage. They also use the transcriptomic characterization of the stromal vascular fraction of the adipose tissue to evaluate molecular and cellular drivers of this sex-specific DIO protection.
Although the …Reviewer #1 (Public Review):
In this manuscript, Li et al characterize sex differences in the impact of macrophage RELMa in protection against diet-induced obesity [DIO]. This is a key area of interest as obesity studies in mice have generally focused exclusively on male animals, as they tend to gain more weight, faster than female mice. The authors use a combination of flow cytometry, adoptive transfer, and single-cell transcriptomics to characterize the mechanism of action for female-specific DIO protection. They identify a potential role for eosinophils in mediating female DIO protection downstream of RELMa production by macrophage. They also use the transcriptomic characterization of the stromal vascular fraction of the adipose tissue to evaluate molecular and cellular drivers of this sex-specific DIO protection.
Although the authors provide solid evidence for many claims in the manuscript, there is generally not enough information about the studies' methods (especially on the computational/data analysis aspects) for a careful evaluation of the result's robustness at this stage. -
Reviewer #2 (Public Review):
In the study by Li et al., the authors hypothesize that RELMa, a macrophage-derived protein, plays a sex-dimorphic role as a protective factor in obesity in females vs males. The authors perform largely in vivo studies utilizing male and female WT and RELMa KO mice on a high-fat diet and perform an in-depth analysis of immune cell composition, gene expression, and single-cell RNA Sequencing. The authors find that WT females are protected from obesity and inflammation vs males, and this protection is lost in female RELMa KO mice. Further analysis by the authors including flow cytometry of the visceral fat SVF in female WT mice showed reduced macrophage infiltration, higher levels of eosinophils, and Th2 cytokine expression compared to WT male mice and female KO mice. The authors show that protection from …
Reviewer #2 (Public Review):
In the study by Li et al., the authors hypothesize that RELMa, a macrophage-derived protein, plays a sex-dimorphic role as a protective factor in obesity in females vs males. The authors perform largely in vivo studies utilizing male and female WT and RELMa KO mice on a high-fat diet and perform an in-depth analysis of immune cell composition, gene expression, and single-cell RNA Sequencing. The authors find that WT females are protected from obesity and inflammation vs males, and this protection is lost in female RELMa KO mice. Further analysis by the authors including flow cytometry of the visceral fat SVF in female WT mice showed reduced macrophage infiltration, higher levels of eosinophils, and Th2 cytokine expression compared to WT male mice and female KO mice. The authors show that protection from obesity and inflammation in female RELMa KO mice can be rescued with an injection of eosinophils and recombinant RELMa. Lastly, the authors use single-cell RNA-Sequencing to further analyze SVF cells in WT and KO male and female mice on a high-fat diet.
Overall, we find that the study represents an important finding in the immunometabolism field showing that RELMa is a key myeloid-derived factor that helps influence the macrophage-eosinophil function in female mice and protects from diet-induced obesity and inflammation in a sexually dimorphic manner. Overall, the study provides strong and convincing data supporting the authors' hypothesis and conclusion.
-
Reviewer #3 (Public Review):
Li, Ruggiero-Ruff et al. examine the role of RELMα, an anti-inflammatory macrophage signature gene, in mediating sex differences in high-fat diet (HFD)-induced obesity in young mice. Specifically, the authors hypothesize that RELMα protects females against HFD-induced obesity. Comparisons between RELMα-knockout (KO) and wildtype (WT) mice of both sexes revealed sex- and RELMα-specific differences in weight gain, immune cell populations, and inflammatory signaling in response to HFD. RELMα-deficiency in females led to increased weight gain, expansion of pro-inflammatory macrophage populations, and eosinophil loss in response to HFD. Female RELMα-deficiency could be rescued by RELMα treatment or eosinophil transfer. Single-cell RNA-sequencing (scRNA-seq) of adipose stromal vascular fraction (SVF) revealed sex- …
Reviewer #3 (Public Review):
Li, Ruggiero-Ruff et al. examine the role of RELMα, an anti-inflammatory macrophage signature gene, in mediating sex differences in high-fat diet (HFD)-induced obesity in young mice. Specifically, the authors hypothesize that RELMα protects females against HFD-induced obesity. Comparisons between RELMα-knockout (KO) and wildtype (WT) mice of both sexes revealed sex- and RELMα-specific differences in weight gain, immune cell populations, and inflammatory signaling in response to HFD. RELMα-deficiency in females led to increased weight gain, expansion of pro-inflammatory macrophage populations, and eosinophil loss in response to HFD. Female RELMα-deficiency could be rescued by RELMα treatment or eosinophil transfer. Single-cell RNA-sequencing (scRNA-seq) of adipose stromal vascular fraction (SVF) revealed sex- and RELMα-dependent differences under HFD conditions and identified potential "pro-obesity" and "anti-obesity" genes in a cell-type-specific manner. Using trajectory analysis, the authors suggest dysregulation of macrophage-to-monocyte transition in RELMα-deficient mice.
The conclusions of this paper are mostly well supported by the data, but some aspects of the statistical and single-cell analyses will need to be corrected, clarified, and extended to enhance the report.
Strengths:
The authors use several orthogonal approaches (i.e., flow cytometry, immunohistochemistry, scRNA-Seq) and models to support their hypotheses.The authors demonstrate that phenotypes observed in HFD-fed females with RELMα-deficiency (i.e., weight gain, loss of eosinophils, a gain of M1 macrophages) can be rescued by RELMα treatment or eosinophil transfer.
The authors recognized the complexity of macrophage activation that is beyond the 'M1/M2' paradigm and informed readers in the introduction as to why this paradigm was used in this study. During the scRNA-seq analyses, the authors further sub-cluster macrophages to include more granularity.
Weaknesses:
There are several instances in the text where the authors claim that there is a significant difference between the two groups, but the statistics for these comparisons are not shown in the figure.It is unfortunate that eosinophils could not be identified in the single-cell analysis since this population of cells was shown to be important in rescuing the RELMα-deficiency in HFD-fed females. The authors should note in the discussion how future scRNA-Seq experiments could overcome this limitation (i.e., enriching immune cells prior to scRNA-Seq).
There are several issues with the scRNA-Seq analysis and interpretation. More details on the steps taken in the single-cell analyses should be included in the methods section. With regards to the 'pseudobulk' analyses presented in Figs. 5-6, several of the differentially expressed genes identified in Fig. 6 are hemoglobin genes (i.e., Hba, Hbb genes). It is not uncommon to filter these genes out of single-cell analysis since their presence usually indicates red blood cell (RBC) contamination (PMID: 31942070, PMID: 35672358). We would recommend assessing RBC contamination as well as removing Fig. 6 from the manuscript and focusing on cell-type-specific analyses. Re-analysis will likely have an impact on the overall conclusions of the study.
Within the text, there are several instances where the authors claim that a pathway is upregulated based on their Gene Ontology (GO) over-representation analysis (ORA). To come to this conclusion, the authors identify genes that are upregulated in one condition and then perform GO-ORA on these genes. However, the authors do not consider negative regulators, whose upregulation would actually decrease the pathway. Authors should either replace their GO-ORA analysis with one that considers the magnitude and direction of differentially expressed genes and provides an activation z-score (i.e., Ingenuity Pathway Analysis) or replace instances of 'upregulated' or 'downregulated' pathways with 'over-represented' pathways.
For Fig.7A, a representative tSNE plot for each group (WT Female, KO Female, WT Male, KO Male) should be shown to ensure there is proper integration of the clusters across groups. There are some instances where the scRNA-Seq data do not appear to be integrated properly (i.e., Supplemental Figure 2C). The authors should explore integration techniques (i.e., Seurat; PMID: 29608179) to correct for potential batch effects within the analysis.
LncRNA Gm47283 is identified as a gene that is differentially expressed by genotype in HFD females (Fig. 7G); however, according to Ensembl this gene is encoded on the Y-chromosome (https://uswest.ensembl.org/Mus_musculus/Gene/Summary?g=ENSMUSG00000096768;r=Y:90796007-90827734). The authors should use the RELMα genotype and sex chromosomally-encoded genes to confirm that their multiplexing was appropriate.
For Fig. 8, samples should be co-clustered and integrated across groups before performing trajectory analysis to allow for direct comparisons between groups.
Since the experiments presented in this report were from young mice using a single diet intervention, the authors should comment on how age and other obesogenic diets may impact the results found here. Also, the authors should expand their discussion as to what upstream regulators (i.e., hormones or genetics) may be driving the sex differences in RELMα expression in response to HFD.
-
