Flexible coding of time or distance in hippocampal cells
Curation statements for this article:-
Curated by eLife

eLife assessment
The manuscript by Abramson and colleagues is a new analysis of previously published data from experiments in which rats ran on a treadmill in either fixed-time or fixed-distance trials. The valuable results provide solid evidence to demonstrate that time and distance cells are more common in fixed-time and fixed-distance trials, respectively. These findings suggest that the hippocampus flexibly shifts between representing variables depending on their relevance.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Analysis of neuronal activity in the hippocampus of behaving animals has revealed cells acting as ‘Time Cells’, which exhibit selective spiking patterns at specific time intervals since a triggering event, and ‘Distance Cells’, which encode the traversal of specific distances. Other neurons exhibit a combination of these features, alongside place selectivity. This study aims to investigate how the task performed by animals during recording sessions influences the formation of these representations. We analyzed data from a treadmill running study conducted by Kraus et al., 2013, in which rats were trained to run at different velocities. The rats were recorded in two trial contexts: a ‘fixed time’ condition, where the animal ran on the treadmill for a predetermined duration before proceeding, and a ‘fixed distance’ condition, where the animal ran a specific distance on the treadmill. Our findings indicate that the type of experimental condition significantly influenced the encoding of hippocampal cells. Specifically, distance-encoding cells dominated in fixed-distance experiments, whereas time-encoding cells dominated in fixed-time experiments. These results underscore the flexible coding capabilities of the hippocampus, which are shaped by over-representation of salient variables associated with reward conditions.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
The idea that because the hippocampal code generates responses that match the most needed variable for each task (time or distance) makes it a predictive code is not fully proved with the analyses provided in the manuscript. For example, in the elapsed time task, there are also place cells and in the fixed-distance travel there are also cells that encode other features. This, rather than a predictive code, can be a regular sample of the environment with an overrepresentation of the more salient variable that animals need to get in order to collect rewards.
We concur with the Reviewer’s reservation. Claims about predictive coding were removed and the following possible account explanation for over-representation was suggested instead:
"These results underscore the flexible coding …
Author Response
Reviewer #1 (Public Review):
The idea that because the hippocampal code generates responses that match the most needed variable for each task (time or distance) makes it a predictive code is not fully proved with the analyses provided in the manuscript. For example, in the elapsed time task, there are also place cells and in the fixed-distance travel there are also cells that encode other features. This, rather than a predictive code, can be a regular sample of the environment with an overrepresentation of the more salient variable that animals need to get in order to collect rewards.
We concur with the Reviewer’s reservation. Claims about predictive coding were removed and the following possible account explanation for over-representation was suggested instead:
"These results underscore the flexible coding capabilities of the hippocampus, which are shaped by over-representation of salient variables associated with reward conditions. " (page 1 line 23, page 4 line 27)
In addition, the analysis provided in the manuscript are rather simple, and better controls could be provided. Improving the analytical quantification of the results is necessary to support the main claim.
We improved the quantification, as suggested below by specific comments of the reviewer.
What is the relationship of each type of cell with the speed of the animal?
The cells were assigned to the different types according to their responses while running across all speeds. However, we checked how the speed of the animal affects the peak firing rate of the cells, for each type of cell. Results of this analysis are presented in Author response image 1. Bars represent maximum firing rate of all cells of a given type across runs with the specified speed range (𝒎𝒆𝒂𝒏 ± 𝑺𝑬𝑴).
Author response image 1.
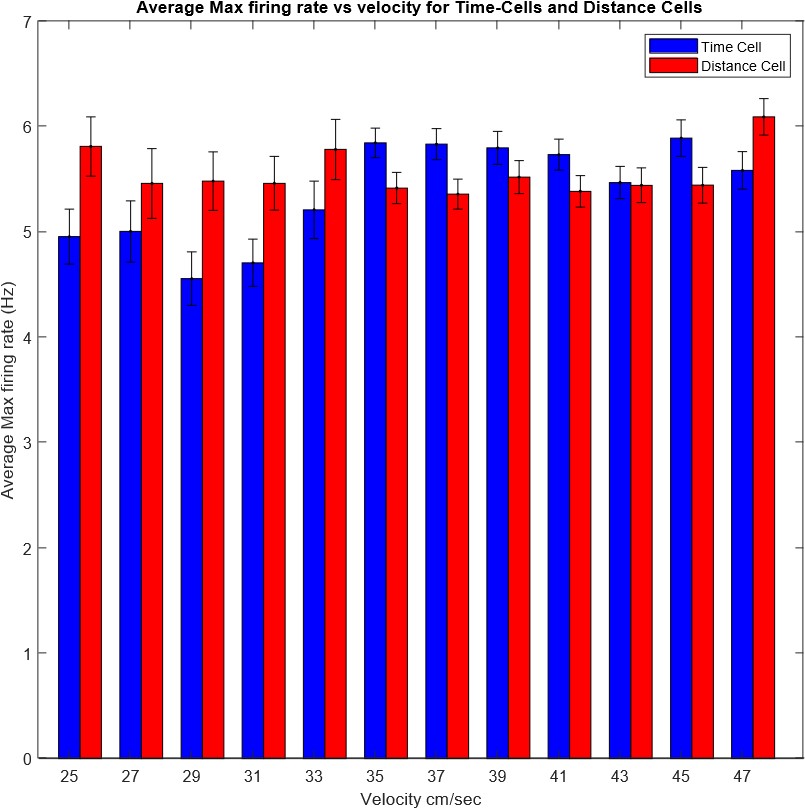
We did not find a significant interaction effect of the speed and the cell-type over the max firing rate (2-way Anova p>0.98).
What is the relationship with the n of trial that the animal has run (first 10 trials, last 10 trials..)?
Some of the animals were subjected to only one type of session. Moreover, they were sometimes trained without recording. Therefore, to answer this question we restricted our analysis to recording sessions where the animal switched from fixed-time to fixed-distance or vice versa. We checked the 20 first runs vs. the last 20 runs (data from 10 runs is not powerful enough for analysis) in See the results in Author response table 1.
Author response table 1.
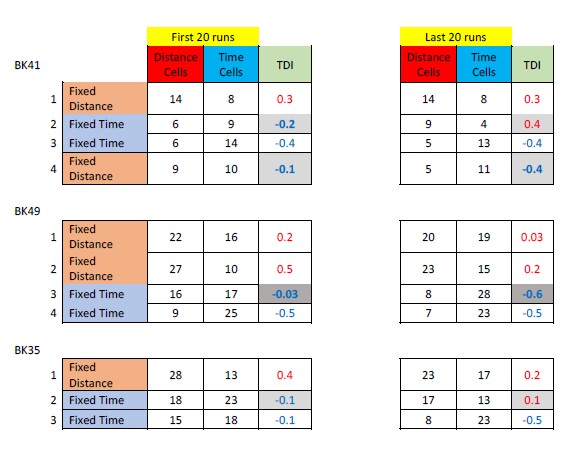
To assess the dynamics of the coding flexibility, we defined the Time-Distance index (TDI), quantifying the balance between the proportion of distance cells and of time cells at a given time. as (NDistanceCells/NTimeCells)/(NDistanceCells+NTimeCells). The is in the range of [0 ,1] if the majority of cells are classified as distance cells, and in the range of [-1, 0] if the majority of cells are classified as time cells. Chi-square testing for differences in proportions did not reveal significant differences (after correction for multiple comparisons).
The shaded boxes in Author response table 1 indicate the sessions which followed a transition between session types
What is the average firing rate of each neuron?
This information was now added to the titles of the panels in Figure 2 and Figure 2-figure supplement 1.
Is there any relationship between intrinsic firing rate and the type of coding that the cell develops in each task?
In Author response image 2 is a comparison of the firing rates of the Time cells vs the Distance cells.
The distributions are similar (p = 0.975 ,and p = 0.675 for peak firing rate and mean firing rate, respectively, Kolmogorov-Smirnov (KS) test).
Author response image 2.
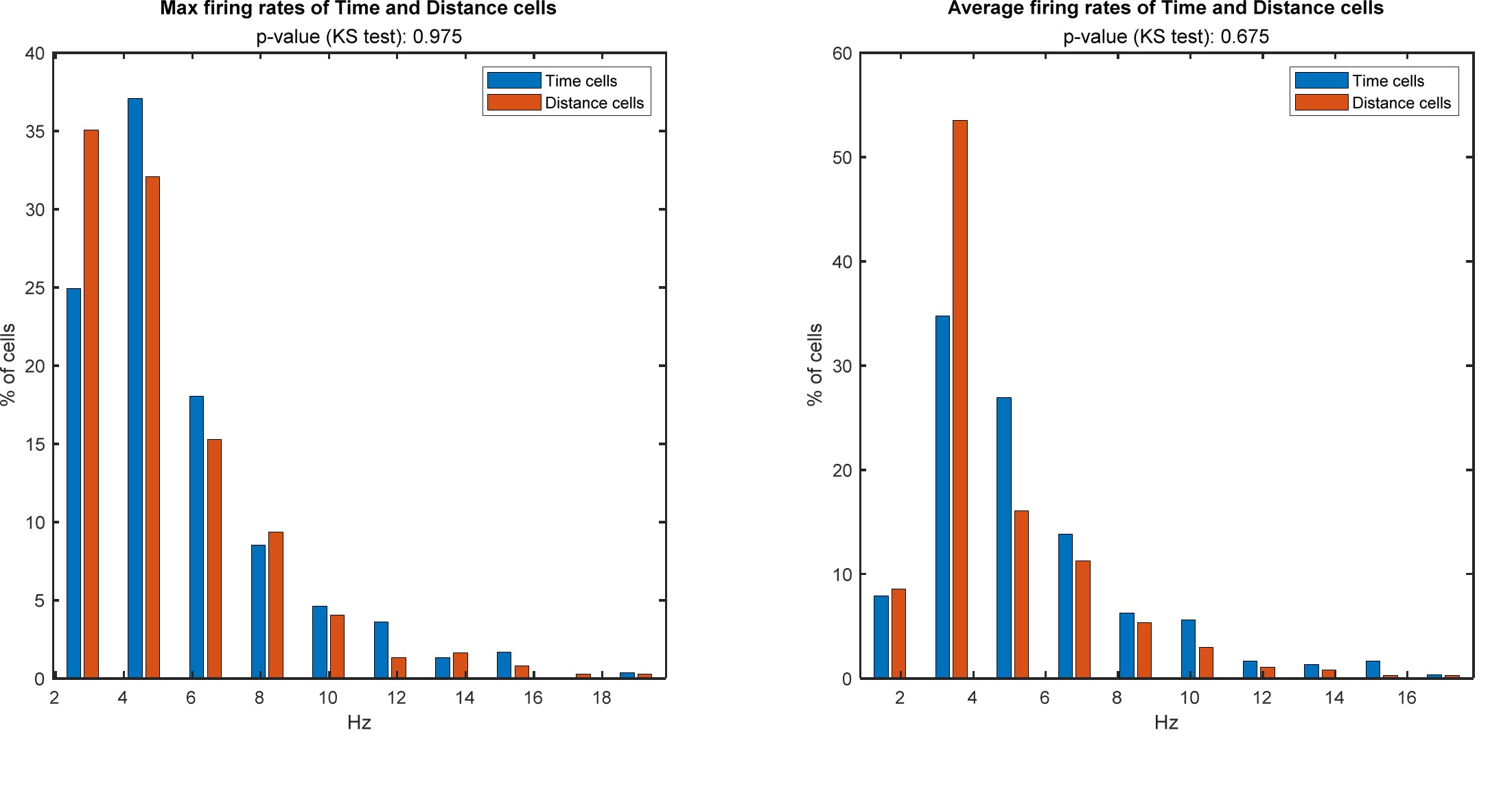
This figure was added to the supplementary figures (figure 3 - figure supplement 3)
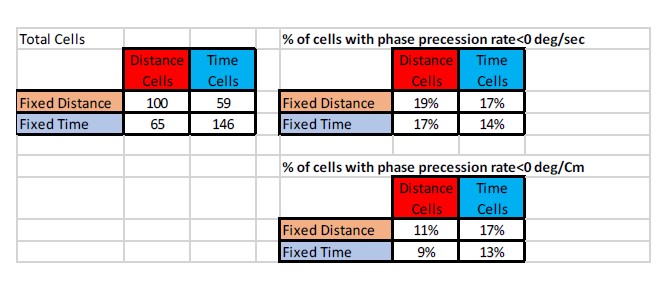
What is the relation of the units of each type with LFP features (theta phase, ripple recruitment)?
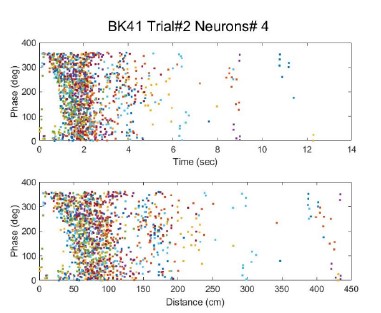
We had LFP recordings for 15 out of 18 sessions. A large proportion of the cells showed phase precession (see Author response table 2). An example is shown in Author response image 3. We could not find a significant relation between phase precession and the cell type or the trial type.
The table on the left shows the total cells analyzed, and on the right we show the percentage of cells that had a significant linear fit of the theta phase within 80% of the field width, when analyzed per time (topright) or per distance (bottom-right). FDist/Ftime are Fixed-distance and fixed-time trials and Dist/Time are the cell type.
We did not identify ripple events during treadmill runs.
Author response table 2
Author response image 3
Reviewer #3 (Public Review):
Weaknesses:
The original study of Kraus et al. consisted of 3 rats for which all sessions, including both training and recording, were of one type. Another 3 rats had a hybrid mixture of distance and time sessions. This is mentioned very briefly in the main text.
It would appear that the theory of reward might lead to different predictions that could be verified by comparing these animals session to session at a finer grain. For example, are there examples of cells switching or transforming their “predictive” representations when a large number of trials in on session type is followed by a large number of trials of the opposite type?
For another example, the transition from training to recording could give similar opportunities. It seems at least possible that ignoring these issues could cause a loss of power.
We could not compare a particular cell for switching between encodings since the different types of trial were performed on different days. As an alternative, we compared the populations of cells within the first 20 vs. last 20 trials in recording sessions where the animal switched from fixed-time to fixed-distance or vice versa (see table below). The “Time-Distance balance index” (TDI) is defined as (#DistanceCells#TimeCells)/(#DistanceCells+#TimeCells) and is ranges between 0 and 1 if the majority of cells are classified as distance cells while between -1 to 0 if the majority of cells are classified as time cells.
In all three animals there seems to be a change between the first 20 runs and last 20 runs of the same session, following a switch between trial types. However, this change is significant and with the expected trend only in one of the animals (BK49, p=0.02, chi-square test).
The grayed boxes in Author response table 1 indicate the sessions which followed a transition between session types
Some circularities in the construction and interpretation of the time-cell and distance-cell classifiers are not clearly addressed. The classifiers currently appear to be fit to predict the type of session a cell’s response patterns are observed within. But it is tautological to use the session type to define the cell type. I sense this is ultimately reasonable because of how the classifier is built, but this concern is not addressed or explained.
We regret that the term ‘classifiers’ was not sufficiently precise. We used this term to describe the metrics designed to express the relation between the firing-time and the velocity, in order to classify cells, rather than classifiers that are fit to predict the type of session. We believe this to be the source of the apparent circularity. To circumvent this confusion, we now replaced all places where the term “classifier” was mentioned, with the term “metric”
-
eLife assessment
The manuscript by Abramson and colleagues is a new analysis of previously published data from experiments in which rats ran on a treadmill in either fixed-time or fixed-distance trials. The valuable results provide solid evidence to demonstrate that time and distance cells are more common in fixed-time and fixed-distance trials, respectively. These findings suggest that the hippocampus flexibly shifts between representing variables depending on their relevance.
-
Reviewer #1 (Public Review):
The idea that because the hippocampal code generates responses that match the most needed variable for each task (time or distance) makes it a predictive code is not fully proved with the analyses provided in the manuscript. For example, in the elapsed time task, there are also place cells and in the fixed-distance travel there are also cells that encode other features. This, rather than a predictive code, can be a regular sample of the environment with an overrepresentation of the more salient variable that animals need to get in order to collect rewards. In addition, the analysis provided in the manuscript are rather simple, and better controls could be provided. Improving the analytical quantification of the results is necessary to support the main claim.
- What is the relationship of each type of cell …
Reviewer #1 (Public Review):
The idea that because the hippocampal code generates responses that match the most needed variable for each task (time or distance) makes it a predictive code is not fully proved with the analyses provided in the manuscript. For example, in the elapsed time task, there are also place cells and in the fixed-distance travel there are also cells that encode other features. This, rather than a predictive code, can be a regular sample of the environment with an overrepresentation of the more salient variable that animals need to get in order to collect rewards. In addition, the analysis provided in the manuscript are rather simple, and better controls could be provided. Improving the analytical quantification of the results is necessary to support the main claim.
- What is the relationship of each type of cell with the speed of the animal?
- What is the relationship with the n of trial that the animal has run (first 10 trials, last 10 trials..)?
- What is the average firing rate of each neuron? Is there any relationship between intrinsic firing rate and the type of coding that the cell develops in each task?
- What is the relation of the units of each type with LFP features (theta phase, ripple recruitment)? -
Reviewer #2 (Public Review):
The manuscript presents a very simple and clear result. It demonstrates that neither place-cell nor time-cell presence is a constant in the rat hippocampus, but that both of these modes of activity are engaged flexibly depending on task demands. This result fits into a growing body of published work showing similar examples of flexibility in hippocampal representations; the authors do a fair job in relating their results to these studies. The innovative aspect of their manuscript is that it specifically addresses place cells and time cells, which have been different, and sometimes confusing, ways of thinking about hippocampal activity. By showing that the hippocampus shifts between distance and time encoding, the authors fit place cells and time cells into a more general framework of flexible representations.
Reviewer #2 (Public Review):
The manuscript presents a very simple and clear result. It demonstrates that neither place-cell nor time-cell presence is a constant in the rat hippocampus, but that both of these modes of activity are engaged flexibly depending on task demands. This result fits into a growing body of published work showing similar examples of flexibility in hippocampal representations; the authors do a fair job in relating their results to these studies. The innovative aspect of their manuscript is that it specifically addresses place cells and time cells, which have been different, and sometimes confusing, ways of thinking about hippocampal activity. By showing that the hippocampus shifts between distance and time encoding, the authors fit place cells and time cells into a more general framework of flexible representations.
The manuscript uses somewhat unusual and not very well-motivated criteria for classifying cells as distance or time cells. To detect the timing of neural activity on each trial, the authors look at the earliest onset of firing prior to the peak. It seems that this method would be highly susceptive to noise, and it is unclear why it would be better than the more standard methods like detecting the actual peak of firing or fitting a stretchable template to the entire firing pattern on each trial. This is a minor weakness of the manuscript, since the main conclusion shouldn't depend on the exact method used to classify cells. The difference between fixed-time and fixed-distance trials reported by the manuscript appears to be large and statistically robust.
-
Reviewer #3 (Public Review):
In some contexts, individual neurons in the hippocampus of rodents, called time cells, can spike selectively after a specific amount of time following a triggering event. Hippocampal neurons can also encode the traversal of a specific amount of distance (for example, running on a treadmill). Some hippocampal neurons also appear to represent mixtures of these features in addition to classical representations of place selectivity. In this manuscript, Abramson et al. hypothesize that the formation of these representations might be influenced by the task which the animal is performing in the context of the recording. To test this hypothesis, they exploit data from a previous maze-running study (Kraus et al., 2013) in which rats were trained to run on a treadmill across several trials of a session at …
Reviewer #3 (Public Review):
In some contexts, individual neurons in the hippocampus of rodents, called time cells, can spike selectively after a specific amount of time following a triggering event. Hippocampal neurons can also encode the traversal of a specific amount of distance (for example, running on a treadmill). Some hippocampal neurons also appear to represent mixtures of these features in addition to classical representations of place selectivity. In this manuscript, Abramson et al. hypothesize that the formation of these representations might be influenced by the task which the animal is performing in the context of the recording. To test this hypothesis, they exploit data from a previous maze-running study (Kraus et al., 2013) in which rats were trained to run on a treadmill across several trials of a session at experimentally-varied velocities. (This study had originally been done to tease apart potential confounds in the questions regarding representations of time versus distance.) In the Kraus et al. study, these walks occurred in one of two contexts or "session types." In a "fixed time" condition, on the other hand, the animal ran on the treadmill for a fixed amount of time before leaving the treadmill. In a "fixed-distance" condition, the animal ran on the treadmill for a "fixed-distance" (in the sense of self-motion). Abramson et al. conjectured that hippocampal pyramidal cells would be biased to represent elapsed time (from entering the treadmill) in the fixed-time condition, whereas they would be biased to represent elapsed distance in the fixed-distance condition. This conjecture appears to be due to the fact that the reward structure of the task motivates the prediction of elapsed time in the fixed time condition, whereas it motivates the prediction of elapsed distance in the fixed distance condition.
To test this hypothesis, the authors use the velocity of the treadmill in each trial to predict the onset of a cell's spiking activity after entering the treadmill. Such predictions would have quite different forms depending on whether the cell's representation correlates with time vs. distance, for example. The authors then use a comparison of the error in each of those two predictors, parametrically formulated, to build a classifier that predicts session type from the spiking onsets of a cell across the trials in that session. The classifier is fit to the Kraus et al. data and optimized to maximize rate of classification as distance cells in the fixed-distance sessions, and minimize rate of classification as time cells in distance sessions. By this metric, they find that 69% of cells in fixed-distance sessions are classified as distance cells, and 68% of cells in the fixed-time sessions are classified as time cells. Applying these results to a parametric hypothesis test, the null hypothesis that session type is independent of cell classifications is strongly rejected. Two other classifiers, based on similar comparisons, found similar results.
The authors conjecture that these findings may be due to the fact that the structure of the task was such that anticipation of reward would depend on "distance" traversed in the fixed-distance sessions, whereas it would depend on time elapsed in the fixed-time sessions. Thus the results are aimed to provide evidence supportive of widely-discussed theories which view the selectivity observed in hippocampal firing patterns as exemplars of predictive coding.
Weaknesses:
The original study of Kraus et al. consisted of 3 rats for which all sessions, including both training and recording, were of one type. Another 3 rats had a hybrid mixture of distance and time sessions. This is mentioned very briefly in the main text. It would appear that the theory of reward might lead to different predictions that could be verified by comparing these animals session to session at a finer grain. For example, are there examples of cells switching or transforming their "predictive" representations when a large number of trials in on session type is followed by a large number of trials of the opposite type? For another example, the transition from training to recording could give similar opportunities. It seems at least possible that ignoring these issues could cause a loss of power.
Some circularities in the construction and interpretation of the time-cell and distance-cell classifiers are not clearly addressed. The classifiers currently appear to be fit to predict the type of session a cell's response patterns are observed within. But it is tautological to use the session type to define the cell type. I sense this is ultimately reasonable because of how the classifier is built, but this concern is not addressed or explained.
Less parametric statistical thinking could be more convincing. Partly this could be a matter of explaining how and why the three classifiers were constructed and their respective scientific motivations. The strong literal finding is the rejection of the hypothesis of independence between cell response properties and session type. A measure of the strength of this effect is missing.
-
