Bridging the gap between presynaptic hair cell function and neural sound encoding
Curation statements for this article:-
Curated by eLife

eLife Assessment
This fundamental study advances substantially our understanding of sound encoding at synapses between single inner hair cells of the mouse cochlea and spiral ganglion neurons. Dual patch-clamp recordings-a technical tour-de force-and careful data analysis provide compelling evidence that the functional heterogeneity of these synapses contributes to the diversity of spontaneous and sound-evoked firing by the neurons. The work will be of broad interest to scientists in the field of auditory neuroscience.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Neural diversity can expand the encoding capacity of a circuitry. A striking example of diverse structure and function is presented by the afferent synapses between inner hair cells (IHCs) and spiral ganglion neurons (SGNs) in the cochlea. Presynaptic active zones at the pillar IHC side activate at lower IHC potentials than those of the modiolar side that have more presynaptic Ca 2+ channels. The postsynaptic SGNs differ in their spontaneous firing rates, sound thresholds, and operating ranges. While a causal relationship between synaptic heterogeneity and neural response diversity seems likely, experimental evidence linking synaptic and SGN physiology has remained difficult to obtain. Here, we aimed at bridging this gap by ex vivo paired recordings of murine IHCs and postsynaptic SGN boutons with stimuli and conditions aimed to mimic those of in vivo SGN characterization. Synapses with high spontaneous rate of release ( SR ) were found predominantly on the pillar side of the IHC. These high SR synapses had larger and more temporally compact spontaneous EPSCs, lower voltage thresholds, tighter coupling of Ca 2+ channels and vesicular release sites, shorter response latencies, and higher initial release rates. This study indicates that synaptic heterogeneity in IHCs directly contributes to the diversity of spontaneous and sound-evoked firing of SGNs.
Article activity feed
-

-

eLife Assessment
This fundamental study advances substantially our understanding of sound encoding at synapses between single inner hair cells of the mouse cochlea and spiral ganglion neurons. Dual patch-clamp recordings-a technical tour-de force-and careful data analysis provide compelling evidence that the functional heterogeneity of these synapses contributes to the diversity of spontaneous and sound-evoked firing by the neurons. The work will be of broad interest to scientists in the field of auditory neuroscience.
-
Reviewer #1 (Public review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size …
Reviewer #1 (Public review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size are 14 vesicles in both high and low SR fibers. 4) Depression is faster in high SR synapses suggesting they have a higher release probability and tighter Ca nanodomain coupling to docked vesicles. 5) Recovery of master-EPSCs from depletion is similar for high and low SR synapses, although there is a slightly faster rate for low SR synapses that have bigger synaptic ribbons, which is very interesting. 6) High SR synapses had larger and more compact (monophasic) sEPSCs, well suited to trigger rapidly and faithfully spikes. 7) High SR synapses exhibit lower voltage (~sound pressure in vivo) dependent thresholds of exocytosis.
Great care was taken to use physiological external pH buffers and physiological external Ca concentrations. Paired recordings were also performed at higher temperatures with IHCs at physiological resting membrane potentials and in more mature animals than previously done for paired recordings. This is extremely challenging because it becomes increasingly difficult to visualize bouton terminals when myelination becomes more prominent in the cochlear afferents. In addition, perforated patch recordings were used in the IHC to preserve its intracellular milieu intact and thus extend the viability of the IHCs. The experiments are tour-de-force and reveal several novel aspects of IHC ribbon synapses. The data set is rich and extensive. The analysis is detailed and compelling.
-
Reviewer #2 (Public review):
Summary:
The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over years. However, the mechanisms that allow the hair cells-SGN …
Reviewer #2 (Public review):
Summary:
The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over years. However, the mechanisms that allow the hair cells-SGN synapses to drive these behaviors are not fully understood.
The level of experimental complexity described in this manuscript is unparalleled, producing data that is hardly found elsewhere. The authors provide strong proof for heterogeneity in transmitter release thresholds at individual synapses and they do so in an extremely complex experimental settings. In addition, the authors found other specific differences such as in synaptic latency and max EPSCs. A reasonable effort is put in bridging these observations with those extensively reported in in vivo SGNs recordings. Similarities are many and differences are not particularly worrying as experimental conditions cannot be perfectly matched, despite the authors' efforts in minimizing them.
-
Reviewer #3 (Public review):
Summary:
The manuscript by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these …
Reviewer #3 (Public review):
Summary:
The manuscript by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these experiments a real tour de force. The data is carefully analyzed and presented, and the results are convincing. In particular, the authors demonstrate that post-synaptic features that contribute to the spontaneous rate (SR) of predominantly monophasic post-synaptic currents (PSCs), shorter EPSC latency, and higher PSC rates are directly paired with pre-synaptic features such as a lower IHC voltage activation and tighter calcium channel coupling for release to give a higher probability of release and subsequent increase in synaptic depression. Importantly, IHCs paired with Low and High SR afferent fibers had the same total calcium currents, indicating that the same IHC can connect to both low and high SR fibers. These fibers also followed expected organizational patterns, with high SR fibers primarily contacting the pillar IHC face and low SR fibers primarily contacting the modiolar face. The authors also use in vivo-like stimulation paradigms to show different RRP and release dynamics that are similar to results from SGN in vivo recordings. Overall, this work systematically examines many features giving rise to specializations and diversity of SGN neurons.
-
Author response:
The following is the authors’ response to the previous reviews.
Reviewer #2 (Recommendations for the authors):
Discussion, page 28. The argument that the authors put forward justifying the (small) size of the spontaneous EPSCs seems reasonable. Nonetheless, it would be good to have an amplitude distribution constructed with voltage-evoked EPSCs to compare with that of spontaneous EPSCs. Not the large initial EPSC, obtained upon IHC depolarization but rather EPSCs occurring later during the longer pulses (figure 4). The authors made the claim that upon IHC depolarization, EPSCs sizes increased, but this is not backed with data.
Following the reviewer recommendation, we have analyzed the voltage-evoked EPSCs occurring during the last 20 ms of the Masker stimulus. We compared the cumulative distribution of the amplitude …
Author response:
The following is the authors’ response to the previous reviews.
Reviewer #2 (Recommendations for the authors):
Discussion, page 28. The argument that the authors put forward justifying the (small) size of the spontaneous EPSCs seems reasonable. Nonetheless, it would be good to have an amplitude distribution constructed with voltage-evoked EPSCs to compare with that of spontaneous EPSCs. Not the large initial EPSC, obtained upon IHC depolarization but rather EPSCs occurring later during the longer pulses (figure 4). The authors made the claim that upon IHC depolarization, EPSCs sizes increased, but this is not backed with data.
Following the reviewer recommendation, we have analyzed the voltage-evoked EPSCs occurring during the last 20 ms of the Masker stimulus. We compared the cumulative distribution of the amplitude of these eEPSCs to the cumulative distribution of the amplitude of the sEPSCs (Figure 1-figure supplement 1, panel G) from the same synapses. The two distributions are significantly different (p < 0.0001, Kolmogorov-Smirnov test), with evoked EPSCs having larger amplitudes (average sEPSC amplitude of -97.28 ± 2.22 pA [median 82.10 pA] vs average eEPSC amplitude of 135.8 ± 3.24 pA [median 120.0 pA]).
-
-
-
eLife Assessment
This fundamental study advances substantially our understanding of sound encoding at synapses between single inner hair cells of the mouse cochlea and spiral ganglion neurons. Dual patch-clamp recordings-a technical tour-de force-and careful data analysis provide compelling evidence that the functional heterogeneity of these synapses contributes to the diversity of spontaneous and sound-evoked firing by the neurons. The work will be of broad interest to scientists in the field of auditory neuroscience.
-
Reviewer #1 (Public review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size …
Reviewer #1 (Public review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size are 14 vesicles in both high and low SR fibers. 4) Depression is faster in high SR synapses suggesting they have a higher release probability and tighter Ca nanodomain coupling to docked vesicles. 5) Recovery of master-EPSCs from depletion is similar for high and low SR synapses, although there is a slightly faster rate for low SR synapses that have bigger synaptic ribbons, which is very interesting. 6) High SR synapses had larger and more compact (monophasic) sEPSCs, well suited to trigger rapidly and faithfully spikes. 7) High SR synapses exhibit lower voltage (~sound pressure in vivo) dependent thresholds of exocytosis.
Great care was taken to use physiological external pH buffers and physiological external Ca concentrations. Paired recordings were also performed at higher temperatures with IHCs at physiological resting membrane potentials and in more mature animals than previously done for paired recordings. This is extremely challenging because it becomes increasingly difficult to visualize bouton terminals when myelination becomes more prominent in the cochlear afferents. In addition, perforated patch recordings were used in the IHC to preserve its intracellular milieu intact and thus extend the viability of the IHCs. The experiments are tour-de-force and reveal several novel aspects of IHC ribbon synapses. The data set is rich and extensive. The analysis is detailed and compelling.
-
Reviewer #2 (Public review):
Summary:
The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over years. However, the mechanisms that allow the hair cells-SGN …
Reviewer #2 (Public review):
Summary:
The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over years. However, the mechanisms that allow the hair cells-SGN synapses to drive these behaviors are not fully understood.
The level of experimental complexity described in this manuscript is unparalleled, producing data that is hardly found elsewhere. The authors provide strong proof for heterogeneity in transmitter release thresholds at individual synapses and they do so in an extremely complex experimental settings. In addition, the authors found other specific differences such as in synaptic latency and max EPSCs. A reasonable effort is put in bridging these observations with those extensively reported in in vivo SGNs recordings. Similarities are many and differences are not particularly worrying as experimental conditions cannot be perfectly matched, despite the authors' efforts in minimizing them.
-
Reviewer #3 (Public review):
Summary:
The manuscript by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these …
Reviewer #3 (Public review):
Summary:
The manuscript by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these experiments a real tour de force. The data is carefully analyzed and presented, and the results are convincing. In particular, the authors demonstrate that post-synaptic features that contribute to the spontaneous rate (SR) of predominantly monophasic post-synaptic currents (PSCs), shorter EPSC latency, and higher PSC rates are directly paired with pre-synaptic features such as a lower IHC voltage activation and tighter calcium channel coupling for release to give a higher probability of release and subsequent increase in synaptic depression. Importantly, IHCs paired with Low and High SR afferent fibers had the same total calcium currents, indicating that the same IHC can connect to both low and high SR fibers. These fibers also followed expected organizational patterns, with high SR fibers primarily contacting the pillar IHC face and low SR fibers primarily contacting the modiolar face. The authors also use in vivo-like stimulation paradigms to show different RRP and release dynamics that are similar to results from SGN in vivo recordings. Overall, this work systematically examines many features giving rise to specializations and diversity of SGN neurons.
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked …
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.
Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size are 14 vesicles in both high and low SR fibers. 4) Depression is faster in high SR synapses suggesting they have a higher release probability and tighter Ca nanodomain coupling to docked vesicles. 5) Recovery of master-EPSCs from depletion is similar for high and low SR synapses, although there is a slightly faster rate for low SR synapses that have bigger synaptic ribbons, which is very interesting. 6) High SR synapses had larger and more compact (monophasic) sEPSCs, well suited to trigger rapidly and faithfully spikes. 7) High SR synapses exhibit lower voltage (~sound pressure in vivo) dependent thresholds of exocytosis.
Strengths:
Great care was taken to use physiological external pH buffers and physiological external Ca concentrations. Paired recordings were also performed at higher temperatures with IHCs at physiological resting membrane potentials and in more mature animals than previously done for paired recordings. This is extremely challenging because it becomes increasingly difficult to visualize bouton terminals when myelination becomes more prominent in the cochlear afferents.
In addition, perforated patch recordings were used in the IHC to preserve its intracellular milieu intact and thus extend the viability of the IHCs. The experiments are tour-de-force and reveal several novel aspects of IHC ribbon synapses. The data set is rich and extensive. The analysis is detailed and compelling.
We would like to thank the reviewer for the appreciation of our work and the comments that helped us to improve our manuscript. We detail our responses to the comments below.
Weaknesses:
(1) Materials and Methods: Please provide whole-cell Rs (series resistance ) and Cm (membrane capacitance) average +/- S.E.M. (or SD) values for IHC and afferent fiber bouton recordings. The Cm values for afferents have been estimated to be about 0.1 pF (Glowatzki and Fuchs, 2002) and it would be interesting to know if there are differences in these numbers for high and low SR afferents. Is it possible to estimate Cm from the capacitative transient time constant? Minimal electronic filtering would be required for that to work, so I realize the authors may not have this data and I also realize that the long cable of the afferents do not allow accurate Cm measurements, but some first order estimate would be very interesting to report, if possible.
In response to the reviewer’s comment, we now added the estimates of series resistance and membrane capacitance for IHC and bouton recordings in Material and Methods and in the Figure 1 – figure supplement 1. Our estimate for bouton Cm is on average 1.7 ± 0.09 pF, a value that compares well to the literature. For example, Glowatzki and Fuchs (2002) provided estimates ranging 0.5-2 pF for recordings from afferent inner hair cell synapses in rats that showed a capacitance transient. In own prior work on afferent inner hair cell synapses of pre-hearing mice, we found estimates of 2.6 ± 0.5 pF (Chapochnikov et al., 2014) and 1.9 ± 0.2 pF (Takago et al., 2019). Keen and Hudspeth (2006) reported capacitances of 1–4 pF for afferent terminals in the bullfrog amphibian papilla. There was no difference in bouton Cm between high SR (1.78 ± 0.19 pF) and low SR synapses (1.68 ± 0.11 pF; p = 0.6575, unpaired t test).
(2) Page 20, 26 and Figure 4: With regard to synaptic delays at auditory hair cell synapses: please see extensive studies done in Figure 11 of Chen and von Gersdorff (JNeurosci., 2019); this showed that synaptic delays are 1.26 ms in adult bullfrog auditory hair cells at 31oC, which is very similar to the High SR fibers (1.19 ms; Fig.4B and page 20). During ongoing depolarizations (e.g. during a sustained sine wave) the synaptic delay can be reduced to just 0.72 ms for probe EPSCs, which is a more usual number for mature fast synapses. This paper should, thus, be cited and briefly discussed in the Discussion. So a significant shortening of delay occurs for the probe response and this is also observed in young rat IHC synapses (see Goutman and Glowatzki, 2011).
We thank the reviewer for this comment. We have analysed the synaptic delay of the probe response and included it in Figure 4 – figure supplement 1. Contrary to the findings from Goutman and Glowatzki (2011) and Chen and von Gersdorff (2019), we did not observe a shortening of the synaptic delay for the probe response compared to the masker response. This difference might arise from the duration of the masker stimulus and/or the IHC holding potential. Synaptic facilitation in hair cells seems to occur only when the RRP is not depleted by the first stimulus (Cho et al., 2011). Our 100 ms masker depolarization from a holding potential of -58 mV effectively depleted the synapse RRP (Figure 4D), while both studies mentioned above used relatively short depolarizations (2 in rat and 20 ms in bullfrog) from a holding potential around -90 mV, which most likely didn’t deplete the RRP. Indeed, when using partially RRP depleting stimuli of 10 ms, Goutman (2011) observed longer synaptic latencies and smaller responses to the second stimulus. We have included this discussion in the last paragraph of the results section.
Additionally, we would also like to note that we referred to the important work on frog hair cell synapses in the manuscript, yet aimed to focus on relating synaptic heterogeneity of mammalian inner hair cell synapses to the functional diversity of type I spiral ganglion neurons that unlike the frog afferents show little branching of their peripheral neurites (in only ~15% of the neurons). We think it will be very interesting to study the aspect of presynaptic heterogeneity in the bullfrog amphibian papilla, but assume that the converging input of several active zones onto a single afferent might provide a different encoding scheme than in the mammalian cochlea.
(3) Gaussian-like (and/or multi-peak) EPSC amplitude distributions were obtained in more mature rat IHCs by Grant et al. (see their Figure 4G; JNeurosci. 2010; postnatal day 19-21). The putative single quanta peak was at 50 pA and the main peak was at 375 pA. The large mean suggests a low CV (probably < 0.4). However, Fig. 2F shows a mean of about 100 pA and CV = 0.7 for spontaneous EPSCs. This major difference deserves some more discussion. I suppose that one possible explanation may be that the current paper holds the IHC membrane potential fixed at -58 mV, whereas Grant et al. (2010) did not control the IHC membrane potential and spontaneous fluctuations in the Vm may have depolarized the IHC, thus producing larger evoked EPSCs that are triggered by Ca channel openings. Some discussion that compares these differences and possible explanations would be quite useful for the readers.
We understand the reviewer’s concern. We have now included the amplitude distribution of sEPSCs recorded from 12 boutons without patch-clamping the IHC (Figure 2–figure supplement 1, panel A). The rest of the recording conditions (i.e., artificial perilymph-like solution, physiological temperature and age) were identical to the conditions used for the paired recordings. Both the range of spontaneous rate (0 up to 16.33 sEPSC/s) and the amplitude distribution (peak at -40 pA and CV of 0.66) were comparable to the values we obtained when clamping the IHC resting potential at -58 mV. In addition, for two of our pairs, we established the bouton recording first, measured the spontaneous release, then established the perforated patch-clamp of the IHC and measured the spontaneous release again with IHC held at -58 mV. For pair #l300321_1, the SR before clamping the IHC was 0.0125 sEPSC/s, with a maximal AmpsEPSC of -110 pA (avg. -52 pA). The SR while holding the IHC at -58 mV was 0.36 sEPSCs/s, with a maximal AmpsEPSC of -140 pA (avg. -46 pA). For pair #l200522_2, the SR changed from 0.07 sEPSC/s to 0. The maximal AmpsEPSC before clamping the IHC was -70 pA (avg. -31 pA). Overall, our data recorded without controlling the IHC argues against the resting potential of -58 mV as a major source of differences in EPSC rate and amplitudes compared to previous studies.
Nonetheless, it is important to note that the experimental conditions used in our study differ from previous reports in several aspects. Our extracellular solution contains the physiological pH buffer bicarbonate instead of the fast buffer HEPES, as well as TEA and Cs+ for proper isolation of the Ca2+ currents. Both pH and potassium channel blockers can alter the excitability of the cell and, consequently, the spontaneous and evoked release. For instance, despite maintaining a similar extracellular pH (7.3 to 7.4), the choice of bicarbonate or HEPES for the extracellular solution can influence differently the regulation of the intracellular pH of the cell (Michl et al., 2019). Indeed, the activity of ion channels and receptors (e.g., AMPAR), and the resting potential can change depending on the extracellular buffer used (Hare and Owen, 1998, Vincent et al., 2019, Cho and von Gersdorff, 2014; and review Sinning and Hübner, 2013). Additionally, the animal model and the age range could be a source of difference. In rats, the EPSC amplitude distribution seems to change with maturation but not with K+ stimulation (Grant et al., 2010) or voltage depolarizations (Goutman and Glowatzki, 2007). This however does not seem to be the case for afferent boutons recorded from mice. In resting conditions (i.e. 5.8 mM extracellular K+), average EPSC amplitudes are around -100 to -150 pA for both prehearing (Chapochnikov et al., 2014) and hearing mice (Niwa et al., 2021 and the present study). Upon stimulation (40 mM K+ or voltage depolarizations), the mean EPSC amplitude does not change in prehearing mice (Jing et al., 2013; Takaba et al., 2019), but it significantly increases in hearing mice (Niwa et al., 2021 and the present study). In p20 and p30 mice, the mean EPSC amplitude was predominantly below -100 pA at rest and only increased above -100 pA after stimulation with 40 mM K+ (Niwa et al., 2021). Similarly, our reported avg. AmpsEPSC is below -150 pA, while the evoked EPSCs reached average amplitudes above -200 pA (Figure 1–figure supplement 1, panel F and Figure 4 – figure supplement 1, panel F).
We have included the aforementioned points in the discussion under the section "Diversity of spontaneous release and their topographical segregation”.
Reviewer #2 (Public Review):
Summary:
The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over the years. However, the mechanisms that allow the hair cells-SGN synapses to drive these behaviors are not fully understood.
Strengths:
The level of experimental complexity described in this manuscript is unparalleled, producing data that is hardly found elsewhere. The authors provide strong proof for heterogeneity in transmitter release thresholds at individual synapses and they do so in extremely complex experimental settings. In addition, the authors found other specific differences such as in synaptic latency and max EPSCs. A reasonable effort is put into bridging these observations with those extensively reported in in vivo SGNs recordings. Similarities are many and differences are not particularly worrying as experimental conditions cannot be perfectly matched, despite the authors' efforts in minimizing them.
We would like to thank the reviewer for the appreciation of our work and the comments that helped us to improve our manuscript. We detail our responses to the comments below.
Weaknesses:
Some concern surges in relation to mismatches with previous reports of IHC-SGN synapses function. EPSCs at these synapses present a peculiar distribution of amplitudes, shapes, and rates. These characteristics are well-established and some do not seem to be paralleled in this study. Here, amplitude distributions are drastically shifted to smaller values, and rates of events are very low, all compared with previous evidence. The reasons for these discrepancies are unclear. The rate at which spontaneous EPSCs appear is an especially sensitive matter. A great part of the conclusions relies on the definition of which of the SGNs (or should say synapses) belong to the low end and which to the high end in the spectrum of spontaneous rates. The data presented by the authors seem a bit off and the criteria used to classify recordings are not well justified. The authors should clarify the origin of these differences since they do not seem to come from obvious reasons such as animal ages, recording techniques, mouse strain, or even species.
We understand the reviewer’s concern. We have now included the amplitude distribution of sEPSCs recorded from 12 boutons without patch-clamping the IHC (Figure 2–figure supplement 1, panel A). The rest of the recording conditions (i.e., artificial perilymph-like solution, physiological temperature and age) were identical to the conditions used for the paired recordings. Both the range of spontaneous rate (0 up to 16.33 sEPSC/s) and the amplitude distribution (peak at -40 pA and CV of 0.66) were comparable to the values we obtained when clamping the IHC resting potential at -58 mV. In addition, for two of our pairs, we established the bouton recording first, measured the spontaneous release, then established the perforated patch-clamp of the IHC and measured the spontaneous release again with IHC held at -58 mV. For pair #l300321_1, the SR before clamping the IHC was 0.0125 sEPSC/s, with a maximal AmpsEPSC of -110 pA (avg. -52 pA). The SR while holding the IHC at -58 mV was 0.36 sEPSCs/s, with a maximal AmpsEPSC of -140 pA (avg. -46 pA). For pair #l200522_2, the SR changed from 0.07 sEPSC/s to 0. The maximal AmpsEPSC before clamping the IHC was -70 pA (avg. -31 pA). Overall, our data recorded without controlling the IHC argues against the resting potential of -58 mV as a major source of differences in EPSC rate and amplitudes compared to previous studies.
Additionally, as noted on the section “Diversity of spontaneous release and their topographical segregation”, our SR values also agree with the range of 0.1 – 16.42 spikes/s reported by Wu et al., (2016) using loose patch recordings from p15-p17 rats. 90% of the paired recordings (and 60% of the bouton recordings) of our dataset were obtained from mice between p14-p17, where spontaneous activity is still low compared to older age groups (p19-p21: 0 – 44.22 spikes/s; p29p32: 0.11 – 54.9 spikes/s Wu et al., 2016; p28: 0 – 47.94 spikes/s, Siebald at al., 2023). There are two additional aspects to consider: i) about 40% of the SGN spikes seem to be generated intrinsically (not activated by an EPSP, ergo an EPSC) at p15-p18 (Wu et al., 2016); and ii) the presence of a spike or EPSC is the sole determinant of a successful recording when the IHC is not stimulated (either by K+ or voltage), thus, these type of experiments undersample fibers with low SR.
We have included the aforementioned points in the discussion under the section "Diversity of spontaneous release and their topographical segregation”.
Reviewer #3 (Public Review):
Summary:
"Bridging the gap between presynaptic hair cell function and neural sound encoding" by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to the diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Strengths
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these experiments a real tour de force. The data is carefully analyzed and presented, and the results are convincing. In particular, the authors demonstrate that post-synaptic features that contribute to the spontaneous rate (SR) of predominantly monophasic post-synaptic currents (PSCs), shorter EPSC latency, and higher PSC rates are directly paired with pre-synaptic features such as a lower IHC voltage activation and tighter calcium channel coupling for release to give a higher probability of release and subsequent increase in synaptic depression. Importantly, IHCs paired with Low and High SR afferent fibers had the same total calcium currents, indicating that the same IHC can connect to both low and high SR fibers. These fibers also followed expected organizational patterns, with high SR fibers primarily contacting the pillar IHC face and low SR fibers primarily contacting the modiolar face. The authors also use in vivo-like stimulation paradigms to show different RRP and release dynamics that are similar to results from SGN in vivo recordings. Overall, this work systematically examines many features giving rise to specializations and diversity of SGN neurons.
We would like to thank the reviewer for the appreciation of our work and the comments that helped us to improve our manuscript. We detail our responses to the comments below.
Weaknesses / Comments / edits:
(1) The careful analysis of calcium coupling and EPSC metrics is especially nice. Can the authors speculate as to why different synapses (likely in the same IHC) would have different calcium cooperativity?
The finding of different apparent Ca2+ cooperativities among IHC synapses is intriguing. Paired pre- and postsynaptic patch-clamp recordings (this work and (Jaime Tobón and Moser, 2023)) and single synapse imaging of presynaptic Ca2+ signals and glutamate release (Özçete and Moser, 2021) jointly support this notion. Both methodologies complement each other. Imaging allows to assess the presynaptic Ca2+ of the specific synapse, while in paired recordings release is related to the whole cell Ca2+ influx. Paired recordings, on the other hand, provide the sensitivity and temporal resolution to assess the initial release rate with short stimuli (2 to 10 ms), which avoids an impact of RRP depletion and ongoing SV replenishment that needs to be considered for the longer stimuli used in imaging (50 ms). Both approaches agree on the finding of tighter coupling of Ca2+ channels and release sites (i.e., lower apparent Ca2+ cooperativity during depolarization within the range of receptor potentials) at pillar synapses. Moreover, the present study took advantage of recording individual release events [which was not achieved by imaging] and further supported the hypothesis that high SR SGNs receive input from active zones with tighter coupling than low SR SGNs. However, our two non-overlapping data sets for paired patch-clamp recordings (this work and (Jaime Tobón and Moser, 2023)) found a narrower range of apparent Ca2+ cooperativities compared to results from single synapse imaging (Özçete and Moser, 2021). This might reflect the technical differences described above. Future studies, potentially combining paired patch-clamp recordings with imaging of presynaptic Ca2+ signals will be needed to scrutinize this aspect.
We think that the different Ca2+ cooperativities reflect subtle differences in the topography of presynaptic Ca2+ channels and vesicular release sites at the specific IHC active zones. The work of Özçete and Moser (2021) indicated that indeed, apparent Ca2+ cooperativities differ among active zones even within the same inner hair cell. Synaptic heterogeneity within one individual cell can expand its coding capacity. In the case of IHCs, differences in the Ca2+ dependence of synaptic release, in addition to the heterogeneous voltage dependence, appears to diversify the response properties (i.e., synaptic vesicle release probability) of individual synapses to the same stimulus. This is particularly important for sound intensity and temporal coding.
We have included the aforementioned points in the discussion under the section "Candidate mechanisms distinguishing evoked release at low and high SR synapses”.
(2) On the bottom of page 6 it would be helpful to mention earlier how many pillar vs modiolar fibers were recorded from, otherwise the skewness of SRs (figure 2H could be thought to be due to predominantly recordings from modiolar fibers. As is, it reads a bit like a cliff-hanger.
Done!
(3) The contrasts for some of the data could be used to point out that while significant differences occur between low and high SR fibers, some of these differences are no longer apparent when comparing modiolar vs pillar fibers (eg by contrasting Figure 2C and 2K). This can indicate that indeed there are differences between the fiber activity, but that the activity likely exists in a gradient across the hair cell faces. Pointing this out at the top of page 10 (end of the first paragraph) would be helpful, it would make the seemingly contradictory voltage dependence data easier to understand on first read (voltage-dependence of release is significantly different between different SR fibers (figure 3) but is not significantly different between fibers on different HC faces (figure S3).
Done!
(4) It should be acknowledged that although the use of post-hearing animals here (P14-23) ensures that SGN have begun to develop more mature activity patterns (Grant et al 2010), the features of the synapses and SGN activity may not be completely mature (Wu et al 2016 PMID: 27733610). Could this explain some of the 'challenges' (authors' section title) detailed on page 28, first full paragraph?
Done!
(5) In the discussion on page 24, the authors compare their recorded SR of EPSCs to measure values in vivo which are higher. Could this indicate that in vivo, the resting membrane potential of IHCs is more depolarized than is currently used for in vitro cochlear experiments?
That is indeed one possible explanation among others. We have expanded the discussion about the factors that could affect the SR in ex vivo experiments.
(6) The results showing lower calcium cooperativity of high SR fibers are powerful, but do the authors have an explanation for why the calcium cooperativity of < 2 is different from that (m = 3-4) observed in other manuscripts?
We assume this question to potentially result from a misunderstanding. Using membrane capacitance measurements and Ca2+ uncaging, Beutner et al. (2001) reported a high intrinsic Ca2+ cooperativity of inner hair cell exocytosis (m = 4-5). Based on this data, it has been proposed that the binding of 4-5 Ca2+ ions is required to trigger the fusion of a synaptic vesicle in IHCs. However, given the shortcoming of Ca2+ uncaging, we and others aimed to further study this aspect using alternative methods. By varying the current of single Ca2+ channels in apical IHCs of hearing mice, several studies reported a high apparent Ca2+ cooperativity (m = 3-5) that is thought to reflect the high intrinsic cooperativity (Brandt et al., 2005; Wong et al., 2014; Özçete and Moser, 2021; Jaime Tobón and Moser, 2023).
On the other hand, the apparent Ca2+ cooperativity observed upon changing the number of open Ca2+ channels would also reflect the active zone topography (i.e., number and distance of Ca2+ channels to the vesicular release site). In the present study, we used different depolarizations within the range of receptor potentials and found a low apparent Ca2+ cooperativity (m < 2) in 93% of the studied synapses. Other studies in apical IHCs from hearing mice used similar and alternative methods to change the number of open Ca2+ channels and also estimated an apparent cooperativity of < 2 (Brandt et al., 2005; Johnson et al., 2005; Johnson et al., 2007; Wong et al., 2014; Özçete and Moser, 2021; Jaime Tobón and Moser, 2023). The fact that these estimates are smaller than those seen upon changes in single Ca2+ current has been taken to indicate that SV release is governed by one or few Ca2+ channels in nanometer proximity (Ca2+ nanodomain-like control of SV exocytosis), building on classical synapse work (Augustine et al., 1991).
In contrast, comparable recordings from mouse IHCs before the onset of hearing (Wong et al., 2014) revealed more similar apparent Ca2+ cooperativities (m ~3) for both changes in the number of open Ca2+ channels and changes in single Ca2+ channel current. This suggests that IHCs before the onset of hearing employ a Ca2+ microdomain-like control of SV exocytosis in which release is governed by the combined activity of several Ca2+ channels in >100 nm distance to the release site. A Ca2+ microdomain-like control of SV exocytosis was also reported for basocochlear IHCs (Johnson et al., 2017).
Recommendations for the authors:
As explained in the public reviews of Reviewers 1 and 2, some mismatches between the data presented here and previous reports from the literature have been identified. It is recommended that you discuss those mismatches, perhaps in relation to the choice of patchclamping the hair cells at -58mV.
We have addressed this point thoroughly in the revised MS. Please see our response to the public review.
Reviewer #1 (Recommendations For The Authors):
Minor suggestions and corrections:
(1) Figures 3 and 4 show beautiful data with paired recordings. Figure 3 shows 10 ms pulses, whereas Fig. 4 shows 100 ms depolarizing pulses. The example in Fig. 3A shows asynchronous release after Ca channel closure, whereas Fig. 4 does not show this so prominently. Was there quite a bit of variability in the asynchronous release from different cell pairs, or was this correlated with pulse duration?
The asynchronous release is also present after 100 ms depolarizing pulses (please see the updated panel A of Figure 4). However, we have not analysed asynchronous release and think that this would be beyond the scope of the current MS. For clarity, we have added dashed lines in the EPSC traces of Figs. 3 and 4 to indicate the on and off-set of the depolarization.
(2) Differences in apex and basal IHC ribbon synapse nanodomain to microdomain Ca channel coupling to exocytosis-sensor have been reported also for gerbil IHCs (see Johnson et al., JNeurosci., 2017). This may be worth mentioning since it is another indication of major synaptic diversity in the mammalian cochlea, this time from low to frequency-located IHCs.
Done
(3) Page 22: change "hight SR" to "high SR".
Done
(4) Page 27: change "addess" to "addressed".
Done
Reviewer #2 (Recommendations For The Authors):
Major points:
(1) As indicated in methods, recording stretches of 5-10 seconds were used to determine the SR of a given SGN. This seems too short for a reasonable estimate of the SR in these neurons. Also, the reported SRs for these mature mice are not only much lower than those measured in in-vivo SGN extracellular recordings but also compared to those reported in ex-vivo rat recordings. Why this discrepancy? The authors decided to estimate SR by voltage-clamping IHCs at a fixed value of - 58 mV, which they take from Johnson, 2015. I wonder if it is not more reasonable to use a range of IHC holdings and measure SR at those, instead of using a single one. It is hard to visualize a very strong argument for using strictly -58 mV. In addition, mapping out a range of holding potentials could provide additional information on IHCs resting membrane potential in physiological conditions.
Related to this point, considering that SR values found in the ex-vivo preparation are much lower than those described in in-vivo situations, is it fair to use the same 1 sp/s criteria, as in Taberner & Liberman, to segregate low and high? Shouldn't this value be adjusted to the overall lower SR? This criterion is naturally critical for the consequent evaluation of other SGN properties.
Finally, on this same problem of IHC Vh, does -58 mV estimate include the 19 mV liquid junction potential? How does it compare with the activation threshold of calcium influx at modiolar vs pillar synapses (see imaging studies)?
We had proactively discussed the challenges of relating ex vivo and in vivo data in the preprint provided for review. While we consider the outcome of our study helpful for better understanding the relation of afferent synaptic heterogeneity and diverse firing properties of SGNs, we do not claim that the assumptions based on literature (such as on the physiological resting potential) represent ground truth.
When carefully revising the MS, we have expanded on the discussion to address the points raised here, particularly regarding the lower SR and sEPSC amplitudes. As this and the other reviewer commented in the public review, these experiments were hard to achieve and we consider repeating them with a range of IHC holding potentials (then not only for spontaneous rate of transmission, but also for in depth characterization of evoked release) to be beyond the scope of the present study.
We do appreciate the suggestion to adjust the distinction between low and high SR given the overall lower rates. However, we would like to refrain from it, as i) we consider it quite arbitrary to define another criterium and ii) we would like to avoid any apparent cherry-picking bias.
Finally, yes, of course, the -58 mV represent the liquid junction potential corrected holding potential. Our average IHC whole-cell Vhalf ICa (-38.86 mV for high SR and -37.60 mV for low SR) compares well with previous reports of average whole-cell Vhalf ICa (-35.44 mV) and average synaptic Vhalf Rhod-FF (-41.15 mV) (Özçete and Moser, 2021). Additionally, our Vhalf QEPSC distribution (ranging from -53.97 to -31.72 mV) also compares well with the Vhalf iGluSnFR distribution (ranging from -45.25 to -29.86 mV) obtained by imaging of synaptic glutamate release (Özçete and Moser, 2021).
- EPSCs amplitude distributions in Figure 2 seem very different from those reported before by Grant et al., 2010 and Niwa et al., 2021 (even Chapochnikov et al., 2014; although not sure if the animal ages match). The average amplitudes of EPSCs reported here, for either pillar or modiolar SGNs, seem way smaller than those reported previously. The authors should provide a convincing explanation for this critical deviation from the consensual results.
Please refer to our response to the public review (point #3).
- Rise time analysis in Fig. 2 supp 1. The actual values seem too long, again, compared to reported values. Also, what would these differences between modiolar and pillar represent?
Previous reports on mouse, rat, turtle and bullfrog focused mainly on the rise times (or time to peak) of monophasic EPSCs: about 0.39 ms (p8-p11 mouse; Chapochnikov et al., 2014, Takago et al., 2019), 0.33-0.58 ms (p7-p14 rat; Yi at al., 2010, Grant et al., 2010, Glowatzki and Fuchs, 2002), 0.17-0.29 ms (p15-p21 rat; Chapochnikov et al., 2014, Huang and Moser, 2018, Grant et al., 2010), 0.1-0.2 ms (turtle auditory papilla; Schnee et al., 2013) and 0.15-0.2 ms (bullfrog 31ºC and 22ºC; Li et al., 2009, Chen and von Gersdorff, 2019). Regarding multiphasic EPSCs, some studies have reported rise times (or times to peak) of about 1.5 ms (p8-p11 mouse; Takago et al., 2019), 1.1 ms (p8-p11 rat; Grant et al., 2010) and 0.6-0.8 ms (p15-p21 rats; Huang and Moser, 2018, Chapochnikov et al., 2014, Grant et al., 2010). When we factor in the waveform of the sEPSCs, our rise times are comparable to the literature:
Author response table 1.
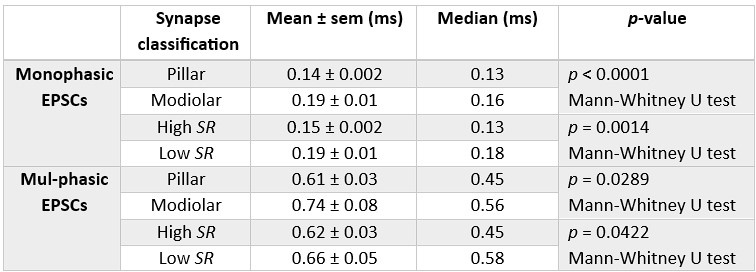
Thus, IHC synapses with higher SR and predominantly located at the pillar side appear to have sEPSCs with faster rise times regardless of their waveform. This might be a consequence of the fusion kinetics of the synaptic vesicles which are tightly influenced by the Ca2+ influx (Huang and Moser, 2018). Additionally, differences in the composition and density of the postsynaptic AMPA receptors could play a role in the rise time of the EPSC (Rubio et al., 2017).
- One of the most impressive observations of the in-vivo SGN physiology is the difference in sound threshold among specific fibers. This can vary over tens of dB of sound pressure levels.
The representation of this phenomenon when using an ex-vivo preparation is not obvious. Overall, it has been reported that IHC Vm is a good proxy for stimulus intensity. Consequently, the authors reported an 'IHC Vm threshold' at the start of SGN synaptic activity for each recording. This can be found in Figure 3 Eii, where values vary between -65 to -30 mV. This is already an important finding. However, the representative traces on panel A only diverge by 5 mV. It would be very interesting to the reader to have represented in the figure recordings that can better illustrate this wide range of values.
We agree with the reviewer regarding the impressive difference in the sound thresholds recorded in vivo. To illustrate better illustrate our findings, we have chosen a different representative trace for the high SR synapse.
- On the masker-probe experiments it would be interesting to look at the synaptic delay of the probe pulses. Are they different between high and low SR synapses?
We have now included the results of the synaptic delay of the probe response (Figure 4– supplementary figure 1). Despite not being statistically significant, the eEPSC probe latency of high SR is on average faster than low SR.
Reviewer #3 (Recommendations For The Authors):
(1) The terms monophasic and compact are used interchangeably. This is fine, but perhaps compact could be defined earlier, otherwise, readers may think that 'compact' means 'short' (as is sometimes euphemistically used to describe short people), which then makes phrasing such as the figure legend for figure 2 a bit confusing. This could be included at first use in a figure as well, in figure 1B where the two types of EPSCs are first shown.
Done, now explained and preferentially used monophasic.
(2) Check for mention of figure panels in the results text - for example, there is no mention in the results text of figure 2A, 2I,
Done
(3) The locations of some of the statistics are inconsistent. This is fine if the authors have a reason for including the stats where they did, but in some cases, the stats are duplicated (for example figure 2J, 2K, 2L, the stats are in both the figure legend and the results text, then check throughout).
Done
(4) The color coding in figure 4 is confusing in panel A - does orange still mean a high SR fiber here? The legend indicates that orange is for EPSCs, but does not specify charge. It could be helpful to show both a high and low SR response, both for EPSCs and for charge.
Thanks for pointing us to this aspect: we have carefully revised the figure and figure legend for clarity. We also included an exemplary response of a low SR synapse in the figure.
-
-
eLife assessment
This fundamental study advances substantially our understanding of sound encoding at synapses between single inner hair cells of the mouse cochlea and spiral ganglion neurons. Dual patch-clamp recordings-a technical tour-de force-and careful data analysis provide compelling evidence that the functional heterogeneity of these synapses contributes to the diversity of spontaneous and sound-evoked firing by the neurons. The work will be of broad interest to scientists in the field of auditory neuroscience.
-
Reviewer #1 (Public Review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size …
Reviewer #1 (Public Review):
Summary:
Tobón and Moser reveal a remarkable amount of presynaptic diversity in the fundamental Ca dependent exocytosis of synaptic vesicles at the afferent fiber bouton synapse onto the pilar or mediolar sides of single inner hair cells of mice. These are landmark findings with profound implications for understanding acoustic signal encoding and presynaptic mechanisms of synaptic diversity at inner hair cell ribbon synapses. The paper will have an immediate and long-lasting impact in the field of auditory neuroscience.Main findings: 1) Synaptic delays and jitter of masker responses are significantly shorter (synaptic delay: 1.19 ms) at high SR fibers (pilar) than at low SR fibers (mediolar; 2.57 ms). 2) Masked evoked EPSC are significantly larger in high SR than in low SR. 3) Quantal content and RRP size are 14 vesicles in both high and low SR fibers. 4) Depression is faster in high SR synapses suggesting they have a higher release probability and tighter Ca nanodomain coupling to docked vesicles. 5) Recovery of master-EPSCs from depletion is similar for high and low SR synapses, although there is a slightly faster rate for low SR synapses that have bigger synaptic ribbons, which is very interesting. 6) High SR synapses had larger and more compact (monophasic) sEPSCs, well suited to trigger rapidly and faithfully spikes. 7) High SR synapses exhibit lower voltage (~sound pressure in vivo) dependent thresholds of exocytosis.
Strengths:
Great care was taken to use physiological external pH buffers and physiological external Ca concentrations. Paired recordings were also performed at higher temperatures with IHCs at physiological resting membrane potentials and in more mature animals than previously done for paired recordings. This is extremely challenging because it becomes increasingly difficult to visualize bouton terminals when myelination becomes more prominent in the cochlear afferents. In addition, perforated patch recordings were used in the IHC to preserve its intracellular milieu intact and thus extend the viability of the IHCs. The experiments are tour-de-force and reveal several novel aspects of IHC ribbon synapses. The data set is rich and extensive. The analysis is detailed and compelling.Weaknesses:
Materials and Methods: Please provide whole-cell Rs (series resistance ) and Cm (membrane capacitance) average +/- S.E.M. (or SD) values for IHC and afferent fiber bouton recordings. The Cm values for afferents have been estimated to be about 0.1 pF (Glowatzki and Fuchs, 2002) and it would be interesting to know if there are differences in these numbers for high and low SR afferents. Is it possible to estimate Cm from the capacitative transient time constant? Minimal electronic filtering would be required for that to work, so I realize the authors may not have this data and I also realize that the long cable of the afferents do not allow accurate Cm measurements, but some first order estimate would be very interesting to report, if possible.
Page 20, 26 and Figure 4: With regard to synaptic delays at auditory hair cell synapses: please see extensive studies done in Figure 11 of Chen and von Gersdorff (JNeurosci., 2019); this showed that synaptic delays are 1.26 ms in adult bullfrog auditory hair cells at 31oC, which is very similar to the High SR fibers (1.19 ms; Fig.4B and page 20). During ongoing depolarizations (e.g. during a sustained sine wave) the synaptic delay can be reduced to just 0.72 ms for probe EPSCs, which is a more usual number for mature fast synapses. This paper should, thus, be cited and briefly discussed in the Discussion. So a significant shortening of delay occurs for the probe response and this is also observed in young rat IHC synapses (see Goutman and Glowatzki, 2011).
Gaussian-like (and/or multi-peak) EPSC amplitude distributions were obtained in more mature rat IHCs by Grant et al. (see their Figure 4G; JNeurosci. 2010; postnatal day 19-21). The putative single quanta peak was at 50 pA and the main peak was at 375 pA. The large mean suggests a low CV (probably < 0.4). However, Fig. 2F shows a mean of about 100 pA and CV = 0.7 for spontaneous EPSCs. This major difference deserves some more discussion. I suppose that one possible explanation may be that the current paper holds the IHC membrane potential fixed at -58 mV, whereas Grant et al. (2010) did not control the IHC membrane potential and spontaneous fluctuations in the Vm may have depolarized the IHC, thus producing larger evoked EPSCs that are triggered by Ca channel openings. Some discussion that compares these differences and possible explanations would be quite useful for the readers.
-
Reviewer #2 (Public Review):
Summary: The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over the years. However, the mechanisms that allow the hair cells-SGN …
Reviewer #2 (Public Review):
Summary: The study by Jaime-Tobon & Moser is a truly major effort to bridge the gap between classical observations on how auditory neurons respond to sounds and the synaptic basis of these phenomena. The so-called spiral ganglion neurons (SGNs) are the primary auditory neurons connecting the brain with hair cells in the cochlea. They all respond to sounds increasing their firing rates, but also present multiple heterogeneities. For instance, some present a low threshold to sound intensity, whereas others have high threshold. This property inversely correlates with the spontaneous rate, i.e., the rate at which each neuron fires in the absence of any acoustic input. These characteristics, along with others, have been studied by many reports over the years. However, the mechanisms that allow the hair cells-SGN synapses to drive these behaviors are not fully understood.
Strengths:
The level of experimental complexity described in this manuscript is unparalleled, producing data that is hardly found elsewhere. The authors provide strong proof for heterogeneity in transmitter release thresholds at individual synapses and they do so in extremely complex experimental settings. In addition, the authors found other specific differences such as in synaptic latency and max EPSCs. A reasonable effort is put into bridging these observations with those extensively reported in in vivo SGNs recordings. Similarities are many and differences are not particularly worrying as experimental conditions cannot be perfectly matched, despite the authors' efforts in minimizing them.Weaknesses:
Some concern surges in relation to mismatches with previous reports of IHC-SGN synapses function. EPSCs at these synapses present a peculiar distribution of amplitudes, shapes, and rates. These characteristics are well-established and some do not seem to be paralleled in this study. Here, amplitude distributions are drastically shifted to smaller values, and rates of events are very low, all compared with previous evidence. The reasons for these discrepancies are unclear. The rate at which spontaneous EPSCs appear is an especially sensitive matter. A great part of the conclusions relies on the definition of which of the SGNs (or should say synapses) belong to the low end and which to the high end in the spectrum of spontaneous rates. The data presented by the authors seem a bit off and the criteria used to classify recordings are not well justified. The authors should clarify the origin of these differences since they do not seem to come from obvious reasons such as animal ages, recording techniques, mouse strain, or even species. -
Reviewer #3 (Public Review):
Summary:
"Bridging the gap between presynaptic hair cell function and neural sound encoding" by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to the diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Strengths
Any of these techniques by themselves are challenging, but the authors do them in pairs, at …Reviewer #3 (Public Review):
Summary:
"Bridging the gap between presynaptic hair cell function and neural sound encoding" by Jaime Tobon and Moser uses patch-clamp electrophysiology in cochlear preparations to probe the pre- and post-synaptic specializations that give rise to the diverse activity of spiral ganglion afferent neurons (SGN). The experiments are quite an achievement! They use paired recordings from pre-synaptic cochlear inner hair cells (IHC) that allow precise control of voltage and therefore calcium influx, with post-synaptic recordings from type I SGN boutons directly opposed to the IHC for both presynaptic control of membrane voltage and post-synaptic measurement of synaptic function with great temporal resolution.
Strengths
Any of these techniques by themselves are challenging, but the authors do them in pairs, at physiological temperatures, and in hearing animals, all of which combined make these experiments a real tour de force. The data is carefully analyzed and presented, and the results are convincing. In particular, the authors demonstrate that post-synaptic features that contribute to the spontaneous rate (SR) of predominantly monophasic post-synaptic currents (PSCs), shorter EPSC latency, and higher PSC rates are directly paired with pre-synaptic features such as a lower IHC voltage activation and tighter calcium channel coupling for release to give a higher probability of release and subsequent increase in synaptic depression. Importantly, IHCs paired with Low and High SR afferent fibers had the same total calcium currents, indicating that the same IHC can connect to both low and high SR fibers. These fibers also followed expected organizational patterns, with high SR fibers primarily contacting the pillar IHC face and low SR fibers primarily contacting the modiolar face. The authors also use in vivo-like stimulation paradigms to show different RRP and release dynamics that are similar to results from SGN in vivo recordings. Overall, this work systematically examines many features giving rise to specializations and diversity of SGN neurons.Weaknesses / Comments / edits:
The careful analysis of calcium coupling and EPSC metrics is especially nice. Can the authors speculate as to why different synapses (likely in the same IHC) would have different calcium cooperativity?
On the bottom of page 6 it would be helpful to mention earlier how many pillar vs modiolar fibers were recorded from, otherwise the skewness of SRs (figure 2H could be thought to be due to predominantly recordings from modiolar fibers. As is, it reads a bit like a cliff-hanger.
The contrasts for some of the data could be used to point out that while significant differences occur between low and high SR fibers, some of these differences are no longer apparent when comparing modiolar vs pillar fibers (eg by contrasting Figure 2C and 2K). This can indicate that indeed there are differences between the fiber activity, but that the activity likely exists in a gradient across the hair cell faces. Pointing this out at the top of page 10 (end of the first paragraph) would be helpful, it would make the seemingly contradictory voltage-dependence data easier to understand on first read (voltage-dependence of release is significantly different between different SR fibers (figure 3) but is not significantly different between fibers on different HC faces (figure S3).
It should be acknowledged that although the use of post-hearing animals here (P14-23) ensures that SGN have begun to develop more mature activity patterns (Grant et al 2010), the features of the synapses and SGN activity may not be completely mature (Wu et al 2016 PMID: 27733610). Could this explain some of the 'challenges' (authors' section title) detailed on page 28, first full paragraph?
In the discussion on page 24, the authors compare their recorded SR of EPSCs to measure values in vivo which are higher. Could this indicate that in vivo, the resting membrane potential of IHCs is more depolarized than is currently used for in vitro cochlear experiments?
The results showing lower calcium cooperativity of high SR fibers are powerful, but do the authors have an explanation for why the calcium cooperativity of < 2 is different from that (m = 3-4) observed in other manuscripts?
-
