The role of conjunctive representations in prioritizing and selecting planned actions
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
Kikumoto and colleagues explore the question of how stimulus- and response-related mental representations are stored and selected in working memory. The authors use a combination of decoding and representational similarity analysis on EEG data to provide evidence for conjunctive representations of action plans. This work would potentially be of great interest to readers in the field of working memory, motor preparation, and selective attention.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #2 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
For flexible goal-directed behavior, prioritizing and selecting a specific action among multiple candidates are often important. Working memory has long been assumed to play a role in prioritization and planning, while bridging cross-temporal contingencies during action selection. However, studies of working memory have mostly focused on memory for single components of an action plan, such as a rule or a stimulus, rather than management of all of these elements during planning. Therefore, it is not known how post-encoding prioritization and selection operate on the entire profile of representations for prospective actions. Here, we assessed how such control processes unfold over action representations, highlighting the role of conjunctive representations that nonlinearly integrate task-relevant features during maintenance and prioritization of action plans. For each trial, participants prepared two independent rule-based actions simultaneously, then they were retro-cued to select one as their response. Prior to the start of the trial, one rule-based action was randomly assigned to be high priority by cueing that it was more likely to be tested. We found that both full action plans were maintained as conjunctive representations during action preparation, regardless of priority. However, during output selection, the conjunctive representation of the high-priority action plan was more enhanced and readily selected as an output. Furthermore, the strength of the high-priority conjunctive representation was associated with behavioral interference when the low-priority action was tested. Thus, multiple alternate upcoming actions were maintained as integrated representations and served as the target of post-encoding attentional selection mechanisms to prioritize and select an action from within working memory.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
The authors ask an interesting question as to whether working memory contains more than one conjunctive representation of multiple task features required for a future response with one of these representations being more likely to become relevant at the time of the response. With RSA the authors use a multivariate approach that seems to become the standard in modern EEG research.
We appreciate the reviewer’s helpful comments on the manuscript and their encouraging comments regarding its potential impact.
I have three major concerns that are currently limiting the meaningfulness of the manuscript: For one, the paradigm uses stimuli with properties that could potentially influence involuntary attention and interfere in a Stroop-like manner with the required responses (i.e., 2 out of 3 cues …
Author Response
Reviewer #1 (Public Review):
The authors ask an interesting question as to whether working memory contains more than one conjunctive representation of multiple task features required for a future response with one of these representations being more likely to become relevant at the time of the response. With RSA the authors use a multivariate approach that seems to become the standard in modern EEG research.
We appreciate the reviewer’s helpful comments on the manuscript and their encouraging comments regarding its potential impact.
I have three major concerns that are currently limiting the meaningfulness of the manuscript: For one, the paradigm uses stimuli with properties that could potentially influence involuntary attention and interfere in a Stroop-like manner with the required responses (i.e., 2 out of 3 cues involve the terms "horizontal" or "vertical" while the stimuli contain horizontal and vertical bars). It is not clear to me whether these potential interactions might bring about what is identified as conjunctive representations or whether they cause these representations to be quite weak.
We agree it is important to rule out any effects of involuntary attention that might have been elicited by our stimulus choices. To address the Reviewer’s concern, we conducted control analyses to test if there was any influence of Stroop-like interference on our measures of behavior or the conjunctive representation. To summarize these analyses (detailed in our responses below and in the supplemental materials), we found no evidence of the effect of compatibility on behavior or on the decoding of conjunctions during either the maintenance or test periods. Furthermore, we found that the decoding of the bar orientation was at chance level during the interval when we observe evidence of the conjunctive representations. Thus, we conclude that the compatibility of the stimuli and the rule did not contribute to the decoding of conjunctive representations or to behavior.
Second, the relatively weak conjunctive representations are making it difficult to interpret null effects such as the absence of certain correlations.
The reviewer is correct that we cannot draw strong conclusions from null findings. We have revised the main text accordingly. In certain cases, we have also included additional analyses. These revisions are described in detail in response the reviewer’s comments below.
Third, if the conjunctive representations truly are reflections of working memory activity, then it would help to include a control condition where memory load is reduced so as to demonstrate that representational strength varies as a function of load. Depending on whether these concerns or some of them can be addressed or ruled out this manuscript has the potential of becoming influential in the field.
This is a clever suggestion for further experimentation. We agree that observing the adverse effect of memory load is one of the robust ways to assess the contributions of working memory system for future studies. However, given that decoding is noisy during the maintenance period (particularly for the low-priority conjunctive representation) even with a relatively low set-size, we expect that in order to further manipulate load, we would need to alter the research design substantially. Thus, as the main goal of the current study is to study prioritization and post-encoding selection of action-related information, we focused on the minimum set-size required for this question (i.e., load 2). However, we now note this load manipulation as a direction for future research in the discussion (pg. 18).
Reviewer #2 (Public Review):
Kikumoto and colleagues investigate the way visual-motor representations are stored in working memory and selected for action based on a retro-cue. They make use of a combination of decoding and RSA to assess at which stages of processing sensory, motor, and conjunctive information (consisting of sensory and motor representations linked via an S- R mapping) are represented in working memory and how these mental representations are related to behavioral performance.
Strengths
This is an elaborate and carefully designed experiment. The authors are able to shed further light on the type of mental representations in working memory that serve as the basis for the selection of relevant information in support of goal- directed actions. This is highly relevant for a better understanding of the role of selective attention and prospective motor representations in working memory. The methods used could provide a good basis for further research in this regard.
We appreciate these helpful comments and the Reviewer’s positive comments on the impact of the work.
Weaknesses
There are important points requiring further clarification, especially regarding the statistical approach and interpretation of results.
- Why is there a conjunction RSA model vector (b4) required, when all information for a response can be achieved by combining the individual stimulus, response, and rule vectors? In Figure 3 it becomes obvious that the conjunction RSA scores do not simply reflect the overlap of the other three vectors. I think it would help the interpretation of results to clearly state why this is not the case.
Thank you for the suggestion, we’ve now added the theoretical background that motivates us to include the RSA model of conjunctive representation (pg. 4 and 5). In particular, several theories of cognitive control have proposed that over the course of action planning, the system assembles an event (task) file which binds all task features at all levels – including the rule (i.e., context), stimulus, and response – into an integrated, conjunctive representation that is essential for an action to be executed (Hommel 2019; Frings et al. 2020). Similarly, neural evidence of non-human primates suggests that cognitive tasks that require context-dependency (e.g., flexible remapping of inputs to different outputs based on the context) recruit nonlinear conjunctive representations (Rigotti et al. 2013; Parthasarathy et al. 2019; Bernardi et al. 2020; Panichello and Buschman, 2021). Supporting these views, we previously observed that conjunctive representations emerge in the human brain during action selection, which uniquely explained behavior such as the costs in transition of actions (Kikumoto & Mayr, 2020; see also Rangel & Hazeltine & Wessel, 2022) or the successful cancelation of actions (Kikumoto & Mayr, 2022). In the current study, by using the same set of RSA models, we attempted to extend the role of conjunctive representations for planning and prioritization of future actions. As in the previous studies (and as noted by the reviewer), the conjunction model makes a unique prediction of the similarity (or dissimilarity) pattern of the decoder outputs: a specific instance of action that is distinct from others actions. This contrasts to other RSA models of low-level features that predict similar patterns of activities for instances that share the same feature (e.g., S-R mappings 1 to 4 share the diagonal rule context). Here, we generally replicate the previous studies showing the unique trajectories of conjunctive representations (Figure 3) and their unique contribution on behavior (Figure 5).
- One of the key findings of this study is the reliable representation of the conjunction information during the preparation phase while there is no comparable effect evident for response representations. This might suggest that two potentially independent conjunctive representations can be activated in working memory and thereby function as the basis for later response selection during the test phase. However, the assumption of the independence of the high and low priority conjunction representations relies only on the observation that there was no statistically reliable correlation between the high and low priority conjunctions in the preparation and test phases. This assumption is not valid because non-significant correlations do not allow any conclusion about the independence of the two processes. A comparable problem appeared regarding the non-significant difference between high and low-priority representations. These results show that it was not possible to prove a difference between these representations prior to the test phase based on the current approach, but they do not unequivocally "suggest that neither action plan was selectively prioritized".
We appreciate this important point. We have taken care in the revision to state that we find evidence of an interference effect for the high-priority action and do not find evidence for such an effect from the low-priority action. Thus, we do not intend to conclude that no such effect could exist. Further, although it is not our intention to draw a strong conclusion from the null effect (i.e., no correlations), we performed an exploratory analysis where we tested the correlation in trials where we observed strong evidence of both conjunctions. Specifically, we binned trials into half within each time point and individual subject and performed the multi-level model analysis using trials where both high and low priority conjunctions were above their medians. Thus, we selected trials in such a way that they are independent of the effect we are testing. The figure below shows the coefficient of associated with low-priority conjunction predicting high-priority conjunction (uncorrected). Even when we focus on trials where both conjunctions are detected (i.e., a high signal-to-noise ratio), we observed no tradeoff. Again, we cannot draw strong conclusions based on the null result of this exploratory analysis. Yet, we can rule out some causes of no correlation between high and low priority conjunctions such as the poor signal-to-noise ratio of the low priority conjunctions. We have further clarified this point in the result (pg. 14).
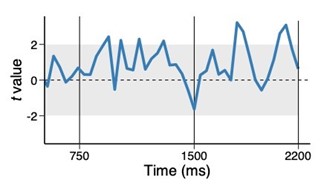
Fig. 1. Trial-to-trial variability between high and low priority conjunctions, using above median trials. The coefficients of the multilevel regression model predicting the variability in trial-to-trial highpriority conjunction by low-priority conjunction.
- The experimental design used does not allow for a clear statement about whether pure motor representations in working memory only emerge with the definition of the response to be executed (test phase). It is not evident from Figure 3 that the increase in the RSA scores strictly follows the onset of the Go stimulus. It is also conceivable that the emergence of a pure motor representation requires a longer processing time. This could only be investigated through temporally varying preparation phases.
We agree with the reviewer. Although we detected no evidence of response representations of both high and low priority action plans during the preparation phase, t(1,23) = -.514, beta = .002, 95% CI [-.010 .006] for high priority; t(1,23) = -1.57, beta = -.008, 95% CI [-.017 .002] for low priority, this may be limited by the relatively short duration of the delay period (750 ms) in this study. However, in our previous studies using a similar paradigm without a delay period (Kikumoto & Mayr, 2020; Kikumoto & Mayr, 2022), response representations were detected less than 300ms after the response was specified, which corresponds to the onset of delay period in this study. Further, participants in the current study were encouraged to prepare responses as early as possible, using adaptive response deadlines and performance-based incentives. Thus, we know of no reason why responses would take longer to prepare in the present study. But we agree that we can’t rule this out. We have added the caveat noted above, as well as this additional context in the discussion (pg. 16-17).
- Inconsistency of statistical approaches: In the methods section, the authors state that they used a cluster-forming threshold and a cluster-significance threshold of p < 0.05. In the results section (Figure 4) a cluster p-value of 0.01 is introduced. Although this concerns different analyses, varying threshold values appear as if they were chosen in favor of significant results. The authors should either proceed consistently here or give very good reasons for varying thresholds.
We thank the reviewer for noting this oversight. All reported significant clusters with cluster P-value were identified using a cluster-forming threshold, p < .05. We fixed the description accordingly.
- Interpretation of results: The significant time window for the high vs. low priority by test-type interaction appeared quite late for the conjunction representation. First, it does not seem reasonable that such an effect appears in a time window overlapping with the motor responses. But more importantly, why should it appear after the respective interaction for the response representation? When keeping in mind that these results are based on a combination of time-frequency analysis, decoding, and RSA (quite many processing steps), I find it hard to really see a consistent pattern in these results that allows for a conclusion about how higher-level conjunctive and motor representations are selected in working memory.
Thank you for raising this important point. First, we fixed reported methodological inconsistencies such as the cluster P-value and cluster-forming threshold). Further, we fully agree that the difference in the time course for the response and conjunctive representations in the low priority, tested condition is unexpected and would complicate the perspective that the conjunctive representation contributes to efficient response selection. However, additional analysis indicates that this apparent pattern in the stimulus locked result is misleading and there is a more parsimonious explanation. First, we wish to caution that the data are relatively noisy and likely are influenced by different frequency bands for different features. Thus, fine-grained temporal differences should be interpreted with caution in the absence of positive statistical evidence of an interaction over time. Indeed, though Figure 4 in the original submission shows a quantitative difference in timing of the interaction effect (priority by test type) across conjunctive representation and response representation, the direct test of this four way interaction [priority x test type x representation type (conjunction vs. response), x time interval (1500 ms to 1850 ms vs. 1850 to 2100 ms)] is not significant, t(1,23) = 1.65, beta = .058, 95% CI [-.012 .015]). The same analysis using response-aligned data is also not significant, t(1,23) = -1.24, beta = -.046, 95% CI [-.128 .028]). These observations were not dependent on the choice of time interval, as other time intervals were also not significant. Therefore, we do not have strong evidence that this is a true timing difference between these conditions and believe this is likely driven by noise.
Further, we believe the apparent late emergence of difference in two conjunctions when the low priority action is tested is more likely due to a slow decline in the strength of the untested high priority conjunction rather than a late emergence of the low priority conjunction. This pattern is clearer when the traces are aligned to the response. The tested low priority conjunction emerges early and is sustained when it is the tested action and declines when it is untested (-226 ms to 86 ms relative to the response onset, cluster-forming threshold, p < .05). These changes eventually resulted in a significant difference in strength between the tested versus untested low priority conjunctions just prior to the commission of the response (Figure 4 - figure supplement 1, the panel on right column of the middle row, the black bars at the top of panel). Importantly, the high priority conjunction also remains active in its untested condition and declines later than the untested low priority conjunction does. Indeed, the untested high priority conjunction does not decline significantly relative to trials when it is tested until after the response is emitted (Figure 4 - figure supplement 1, the panel on right column of the middle row, the red bars at the top of panel). This results in a late emerging interaction effect of the priority and test type, but this is not due to a late emerging low priority conjunctive representation.
In summary, we do not have statistical evidence of a time by effect interaction that allows us to draw strong inferences about timing. Nonetheless, even the patterns we observe are inconsistent with a late emerging low priority conjunctive representation. And if anything, they support a late decline in the untested high priority conjunctive representation. This pattern of the result of the high priority conjunction being sustained until late, even when it is untested, is also notable in light of our observation that the strength of the high priority conjunctive representation interferes behavior when the low priority item is tested, but not vice versa. We now address this point about the timing directly in the results (pg. 15-16) and the discussion (pg. 21), and we include the response locked results in the main text along with the stimulus locked result including exploratory analyses reported here.
Reviewer #3 (Public Review):
This study aims to address the important question of whether working memory can hold multiple conjunctive task representations. The authors combined a retro-cue working memory paradigm with their previous task design that cleverly constructed multiple conjunctive tasks with the same set of stimuli, rules, and responses. They used advanced EEG analytical skills to provide the temporal dynamics of concurrent working memory representation of multiple task representations and task features (e.g., stimulus and responses) and how their representation strength changes as a function of priority and task relevance. The results generally support the authors' conclusion that multiple task representations can be simultaneously manipulated in working memory.
We appreciate these helpful comments, and were pleased that the reviewer shares our view that these results may be broadly impactful.
-
Evaluation Summary:
Kikumoto and colleagues explore the question of how stimulus- and response-related mental representations are stored and selected in working memory. The authors use a combination of decoding and representational similarity analysis on EEG data to provide evidence for conjunctive representations of action plans. This work would potentially be of great interest to readers in the field of working memory, motor preparation, and selective attention.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #2 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
The authors ask an interesting question as to whether working memory contains more than one conjunctive representation of multiple task features required for a future response with one of these representations being more likely to become relevant at the time of the response. With RSA the authors use a multivariate approach that seems to become the standard in modern EEG research.
I have three major concerns that are currently limiting the meaningfulness of the manuscript: For one, the paradigm uses stimuli with properties that could potentially influence involuntary attention and interfere in a Stroop-like manner with the required responses (i.e., 2 out of 3 cues involve the terms "horizontal" or "vertical" while the stimuli contain horizontal and vertical bars). It is not clear to me whether these potential …
Reviewer #1 (Public Review):
The authors ask an interesting question as to whether working memory contains more than one conjunctive representation of multiple task features required for a future response with one of these representations being more likely to become relevant at the time of the response. With RSA the authors use a multivariate approach that seems to become the standard in modern EEG research.
I have three major concerns that are currently limiting the meaningfulness of the manuscript: For one, the paradigm uses stimuli with properties that could potentially influence involuntary attention and interfere in a Stroop-like manner with the required responses (i.e., 2 out of 3 cues involve the terms "horizontal" or "vertical" while the stimuli contain horizontal and vertical bars). It is not clear to me whether these potential interactions might bring about what is identified as conjunctive representations or whether they cause these representations to be quite weak. Second, the relatively weak conjunctive representations are making it difficult to interpret null effects such as the absence of certain correlations. Third, if the conjunctive representations truly are reflections of working memory activity, then it would help to include a control condition where memory load is reduced so as to demonstrate that representational strength varies as a function of load.
Depending on whether these concerns or some of them can be addressed or ruled out this manuscript has the potential of becoming influential in the field.
-
Reviewer #2 (Public Review):
Kikumoto and colleagues investigate the way visual-motor representations are stored in working memory and selected for action based on a retro-cue. They make use of a combination of decoding and RSA to assess at which stages of processing sensory, motor, and conjunctive information (consisting of sensory and motor representations linked via an S-R mapping) are represented in working memory and how these mental representations are related to behavioral performance.
Strengths
This is an elaborate and carefully designed experiment. The authors are able to shed further light on the type of mental representations in working memory that serve as the basis for the selection of relevant information in support of goal-directed actions. This is highly relevant for a better understanding of the role of selective …Reviewer #2 (Public Review):
Kikumoto and colleagues investigate the way visual-motor representations are stored in working memory and selected for action based on a retro-cue. They make use of a combination of decoding and RSA to assess at which stages of processing sensory, motor, and conjunctive information (consisting of sensory and motor representations linked via an S-R mapping) are represented in working memory and how these mental representations are related to behavioral performance.
Strengths
This is an elaborate and carefully designed experiment. The authors are able to shed further light on the type of mental representations in working memory that serve as the basis for the selection of relevant information in support of goal-directed actions. This is highly relevant for a better understanding of the role of selective attention and prospective motor representations in working memory. The methods used could provide a good basis for further research in this regard.Weaknesses
There are important points requiring further clarification, especially regarding the statistical approach and interpretation of results.- Why is there a conjunction RSA model vector (b4) required, when all information for a response can be achieved by combining the individual stimulus, response, and rule vectors? In Figure 3 it becomes obvious that the conjunction RSA scores do not simply reflect the overlap of the other three vectors. I think it would help the interpretation of results to clearly state why this is not the case.
- One of the key findings of this study is the reliable representation of the conjunction information during the preparation phase while there is no comparable effect evident for response representations. This might suggest that two potentially independent conjunctive representations can be activated in working memory and thereby function as the basis for later response selection during the test phase. However, the assumption of the independence of the high and low priority conjunction representations relies only on the observation that there was no statistically reliable correlation between the high and low priority conjunctions in the preparation and test phases. This assumption is not valid because non-significant correlations do not allow any conclusion about the independence of the two processes. A comparable problem appeared regarding the non-significant difference between high and low-priority representations. These results show that it was not possible to prove a difference between these representations prior to the test phase based on the current approach, but they do not unequivocally "suggest that neither action plan was selectively prioritized".
- The experimental design used does not allow for a clear statement about whether pure motor representations in working memory only emerge with the definition of the response to be executed (test phase). It is not evident from Figure 3 that the increase in the RSA scores strictly follows the onset of the Go stimulus. It is also conceivable that the emergence of a pure motor representation requires a longer processing time. This could only be investigated through temporally varying preparation phases.
- Inconsistency of statistical approaches: In the methods section, the authors state that they used a cluster-forming threshold and a cluster-significance threshold of p < 0.05. In the results section (Figure 4) a cluster p-value of 0.01 is introduced. Although this concerns different analyses, varying threshold values appear as if they were chosen in favor of significant results. The authors should either proceed consistently here or give very good reasons for varying thresholds.
- Interpretation of results: The significant time window for the high vs. low priority by test-type interaction appeared quite late for the conjunction representation. First, it does not seem reasonable that such an effect appears in a time window overlapping with the motor responses. But more importantly, why should it appear after the respective interaction for the response representation? When keeping in mind that these results are based on a combination of time-frequency analysis, decoding, and RSA (quite many processing steps), I find it hard to really see a consistent pattern in these results that allows for a conclusion about how higher-level conjunctive and motor representations are selected in working memory.
-
Reviewer #3 (Public Review):
This study aims to address the important question of whether working memory can hold multiple conjunctive task representations. The authors combined a retro-cue working memory paradigm with their previous task design that cleverly constructed multiple conjunctive tasks with the same set of stimuli, rules, and responses. They used advanced EEG analytical skills to provide the temporal dynamics of concurrent working memory representation of multiple task representations and task features (e.g., stimulus and responses) and how their representation strength changes as a function of priority and task relevance. The results generally support the authors' conclusion that multiple task representations can be simultaneously manipulated in working memory.
My only concern is that in Figure 4, the strongest priority by …
Reviewer #3 (Public Review):
This study aims to address the important question of whether working memory can hold multiple conjunctive task representations. The authors combined a retro-cue working memory paradigm with their previous task design that cleverly constructed multiple conjunctive tasks with the same set of stimuli, rules, and responses. They used advanced EEG analytical skills to provide the temporal dynamics of concurrent working memory representation of multiple task representations and task features (e.g., stimulus and responses) and how their representation strength changes as a function of priority and task relevance. The results generally support the authors' conclusion that multiple task representations can be simultaneously manipulated in working memory.
My only concern is that in Figure 4, the strongest priority by task-relevance interaction occurred earlier in the response than the conjunction representation, which seems to be opposite to the assumption that the conjunction representation produces the response and thus requires more discussion on why this is the case. This study expands the working memory research by showing that working memory can simultaneously hold and manipulate multiple task representations. It also provides solid foundation for future work to investigate the control mechanisms on working memory representations of task conjunctions.
-
