RNA localization mechanisms transcend cell morphology
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
Goering and colleagues investigate subcellular RNA localization across different cell types and species. The major insight is that there may be general mechanisms and specifically conserved proteins that regulate RNA localization in diverse cell types and morphologies. This manuscript will be of interest to those studying gene expression and how its regulation occurs within the cell.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
RNA molecules are localized to specific subcellular regions through interactions between RNA regulatory elements and RNA binding proteins (RBPs). Generally, our knowledge of the mechanistic details behind the localization of a given RNA is restricted to a particular cell type. Here, we show that RNA/RBP interactions that regulate RNA localization in one cell type predictably regulate localization in other cell types with vastly different morphologies. To determine transcriptome-wide RNA spatial distributions across the apicobasal axis of human intestinal epithelial cells, we used our recently developed RNA proximity labeling technique, Halo-seq. We found that mRNAs encoding ribosomal proteins (RP mRNAs) were strongly localized to the basal pole of these cells. Using reporter transcripts and single-molecule RNA FISH, we found that pyrimidine-rich motifs in the 5′ UTRs of RP mRNAs were sufficient to drive basal RNA localization. Interestingly, the same motifs were also sufficient to drive RNA localization to the neurites of mouse neuronal cells. In both cell types, the regulatory activity of this motif was dependent on it being in the 5′ UTR of the transcript, was abolished upon perturbation of the RNA-binding protein LARP1, and was reduced upon inhibition of kinesin-1. To extend these findings, we compared subcellular RNAseq data from neuronal and epithelial cells. We found that the basal compartment of epithelial cells and the projections of neuronal cells were enriched for highly similar sets of RNAs, indicating that broadly similar mechanisms may be transporting RNAs to these morphologically distinct locations. These findings identify the first RNA element known to regulate RNA localization across the apicobasal axis of epithelial cells, establish LARP1 as an RNA localization regulator, and demonstrate that RNA localization mechanisms cut across cell morphologies.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
In this manuscript, Goering et al. investigate subcellular RNA localization across different cell types focusing on epithelial cells (mouse C2bbe1 and human HCA-7 enterocyte monolayers, canine MDCK epithelial cells) as well as neuronal cultures (mouse CAD cells). They use their recently established Halo-seq method to investigate transcriptome-wide RNA localization biases in C2bbe1 enterocyte monolayers and find that 5'TOP-motif containing mRNAs, which encode ribosomal proteins (RPs), are enriched on the basal side of these cells. These results are supported by smFISH against endogenous RP-encoding mRNAs (RPL7 and RPS28) as well as Firefly luciferase reporter transcripts with and without mutated 5'TOP sequences. Furthermore, they find that 5'TOP-motifs are not only driving localization to …
Author Response
Reviewer #1 (Public Review):
In this manuscript, Goering et al. investigate subcellular RNA localization across different cell types focusing on epithelial cells (mouse C2bbe1 and human HCA-7 enterocyte monolayers, canine MDCK epithelial cells) as well as neuronal cultures (mouse CAD cells). They use their recently established Halo-seq method to investigate transcriptome-wide RNA localization biases in C2bbe1 enterocyte monolayers and find that 5'TOP-motif containing mRNAs, which encode ribosomal proteins (RPs), are enriched on the basal side of these cells. These results are supported by smFISH against endogenous RP-encoding mRNAs (RPL7 and RPS28) as well as Firefly luciferase reporter transcripts with and without mutated 5'TOP sequences. Furthermore, they find that 5'TOP-motifs are not only driving localization to the basal side of epithelial cells but also to neuronal processes. To investigate the molecular mechanism behind the observed RNA localization biases, they reduce expression of several Larp proteins and find that RNA localization is consistently Larp1-dependent. Additionally, the localization depends on the placement of the TOP sequence in the 5'UTR and not the 3'UTR. To confirm that similar RNA localization biases can be conserved across cell types for other classes of transcripts, they perform similar experiments with a GA-rich element containing Net1 3'UTR transcript, which has previously been shown to exhibit a strong localization bias in several cell types. In order to determine if motor proteins contribute to these RNA distributions, they use motor protein inhibitors to confirm that the localization of individual members of both classes of transcripts, 5'TOP and GA-rich, is kinesin-dependent and that RNA localization to specific subcellular regions is likely to coincide with RNA localization to microtubule plus ends that concentrate in the basal side of epithelial cells as well as in neuronal processes.
In summary, Goering et al. present an interesting study that contributes to our understanding of RNA localization. While RNA localization has predominantly been studied in a single cell type or experimental system, this work looks for commonalities to explain general principles. I believe that this is an important advance, but there are several points that should be addressed.
Comments:
- The Mili lab has previously characterized the localization of ribosomal proteins and NET1 to protrusions (Wang et al, 2017, Moissoglu et al 2019, Crisafis et al., 2020) and the role of kinesins in this localization (Pichon et al, 2021). These papers should be cited and their work discussed. I do not believe this reduces the novelty of this study and supports the generality of the RNA localization patterns to additional cellular locations in other cell types.
This was an unintentional oversight on our part, and we apologize. We have added citations for the mentioned publications and discussed our work in the context of theirs.
- The 5'TOP motif begins with an invariant C nucleotide and mutation of this first nucleotide next to the cap has been shown to reduce translation regulation during mTOR inhibition (Avni et al, 1994 and Biberman et al 1997) and also Lapr1 binding (Lahr et al, 2017). Consequently, it is not clear to me if RPS28 initiates transcription with an A as indicated in Figure 3B. There also seems to be some differences in published CAGE datasets, but this point needs to be clarified. Additionally, it is not clear to me how the 5'TOP Firefly luciferase reporters were generated and if the transcription start site and exact 5'-ends of these constructs were determined. This is again essential to determine if it is a pyrimidine sequence in the 5'UTR that is important for localization or the 5'TOP motif and if Larp1 is directly regulating the localization by binding to the 5'TOP motif or if the effect they observe is indirect (e.g. is Larp1 also basally localized?). It should also be noted that Larp1 has been suggested to bind pyrimidine-rich sequences in the 5'UTR that are not next to the cap, but the details of this interaction are less clear (Al-Ashtal et al, 2021)
We did not fully appreciate the subtleties related to TOP motif location when we submitted this manuscript, so we thank the reviewer for pointing them out.
We also analyzed public CAGE datasets (Andersson et al, 2014 Nat Comm) and found that the start sites for both RPL7 and RPS28 were quite variable within a window of several nucleotides (as is the case for the vast majority of genes), suggesting that a substantial fraction of both do not begin with pyrimidines (Reviewer Figure 1). Yet, by smFISH, endogenous RPL7 and RPS28 are clearly basally/neurite localized (see new figure 3C).
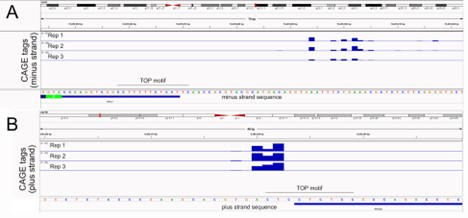
Reviewer Figure 1. Analysis of transcription start sites for RPL7 (A) and RPS28 (B) using CAGE data (Andersson et al, 2014 Nat Comm). Both genes show a window of transcription start sites upstream of current gene models (blue bars at bottom).
A more detailed analysis of our PRRE-containing reporter transcripts led us to find that in these reporters, the pyrimidine-rich element was approximately 90 nucleotides into the body of the 5’ UTR. Yet these reporters are also basally/neurite localized. The organization of the PRRE-containing reporters is now more clearly shown in an updated figure 3D.
From these results, it would seem that the pyrimidine-rich element need not be next to the 5’ cap in order to regulate RNA localization. To generalize this result, we first used previously identified 5’ UTR pyrimidine-rich elements that had been found to regulate translation in an mTOR-dependent manner (Hsieh et al 2012). We found that, as a class, RNAs containing these motifs were similarly basally/neurite localized as RP mRNAs. These results are presented in figures 3A and 3I.
We then asked if the position of the pyrimidine-rich element within the 5’ UTR of these RNAs was related to their localization. We found no relationship between element position and transcript localization as elements within the bodies of 5’ UTRs were seemingly just as able to promote basal/neurite localization as elements immediately next to the 5’ cap. These results are presented in figures 3B and 3J.
To further confirm that pyrimidine-rich elements need not be immediately next to the 5’ cap, we redesigned our RPL7-derived reporter transcripts such that the pyrimidine-rich motif was immediately adjacent to the 5’ cap. This was possible because the reporter uses a CMV promoter that reliably starts transcription at a known nucleotide. We then compared the localization of this reporter (called “RPL7 True TOP”) to our previous reporter in which the pyrimidine-rich element was ~90 nt into the 5’ UTR (called “RPL7 PRRE”) (Reviewer Figure 2). As with the PRRE reporter, the True TOP reporter drove RNA localization in both epithelial and neuronal cells while purine-containing mutant versions of the True TOP reporter did not (Reviewer Figure 2A-D). In the epithelial cells, the True TOP was modestly but significantly better at driving basal RNA localization than the PRRE (Reviewer Figure 2E) while in neuronal cells the True TOPs were modestly but insignificantly better. Again, this suggests that pyrimidine-rich motifs need not be immediately cap-adjacent in order to regulate RNA localization.
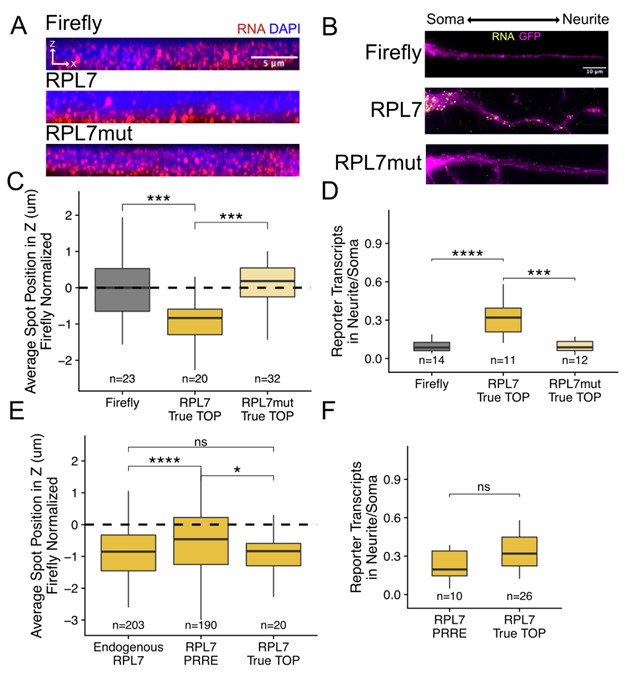
Reviewer Figure 2. Experimental confirmation that pyrimidine-rich motif location within 5’ UTRs is not critical for RNA localization. (A) RPL7 True TOP smFISH in epithelial cells. (B) RPL7 True TOP smFISH in neuronal cells. (C) Quantification of epithelial cell smFISH in A. (D) Quantification of neuronal cell smFISH in D. (E) Comparison of the location in epithelial cells of endogenous RPL7 transcripts, RPL7 PRRE reporter transcripts, and PRL7 True TOP reporter transcripts. (F) Comparison of the neurite-enrichment of RPL7 PRRE reporters and RPL7 True TOP reporters. In C-F, the number of cells included in each analysis is shown.
In response to the point about whether the localization results are direct effects of LARP1, we did not assay the binding of LARP1 to our PRRE-containing reporters, so we cannot say for sure. However, given that PRRE-dependent localization required LARP1 and there is much evidence about LARP1 binding pyrimidine-rich elements (including those that are not cap-proximal as the reviewer notes), we believe this to be the most likely explanation.
It should also be noted here that while pyrimidine-rich motif position within the 5’ UTR may not matter, its location within the transcript does. PRREs located within 3’ UTRs were unable to direct RNA localization (Figure 5).
- In figure 1A, they indicate that mRNA stability can contribute to RNA localization, but this point is never discussed. This may be important to their work since Larp1 has also been found to impact mRNA half-lives (Aoki et al, 2013 and Mattijssen et al 2020, Al-Ashtal et al 2021). Is it possible the effect they see when Larp1 is depleted comes from decreased stability?
We found that PRRE-containing reporter transcripts were generally less abundant than their mutant counterparts in C2bbe1, HCA7, and MDCK cells (figure 3 – figure supplements 5, 6, and 8) although the effect was not consistent in mouse neuronal cells (figure 3 – figure supplement 13).
However, we don’t think it is likely that the changes in localization are due to stability changes. This abundance effect did not seem to be LARP1-dependent as both PRRE-containing and PRRE-mutant reporters were generally more expressed in LARP1-rescue epithelial cells than in LARP1 KO cells (figure 4 – figure supplement 9).
It should be noted here that we are not ever actually measuring transcript stability but rather steady state abundances. It cannot therefore be ruled out that LARP1 is regulating the stability of our PRRE reporters. Given, though, that their localization was dependent on kinesin activity (figures 7F, 7G), we believe the most likely explanation for the localization effects is active transport.
- Also Moor et al, 2017 saw that feeding cycles changed the localization of 5'TOP mRNAs. Similarly, does mTOR inhibition or activation or simply active translation alter the localization patterns they observe? Further evidence for dynamic regulation of RNA localization would strengthen this paper
We are very interested in this and have begun exploring it. We have data suggesting that PRREs also mediate the feeding cycle-dependent relocalization of RP mRNAs. As the reviewer says, we think this leads to a very attractive model involving mTOR, and we are currently working to test this model. However, we don’t have the room to include those results in this manuscript and would instead prefer to include them in a later manuscript that focuses on nutrient-induced dynamic relocalization.
- For smFISH quantification, is every mRNA treated as an independent measurement so that the statistics are calculated on hundreds of mRNAs? Large sample sizes can give significant p-values but have very small differences as observe for Firefly vs. OSBPL3 localization. Since determining the biological interpretation of effect size is not always clear, I would suggest plotting RNA position per cell or only treat biological replicates as independent measurements to determine statistical significance. This should also be done for other smFISH comparisons
This is a good suggestion, and we agree that using individual puncta as independent observations will artificially inflate the statistical power in the experiment. To remedy this in the epithelial cell images, we first reanalyzed the smFISH images using each of the following as a unique observation: the mean location of all smFISH puncta in one cell, the mean location of all puncta in a field of view, and the mean location of all puncta in one coverslip. With each metric, the results we observed were very similar (Reviewer Figure 3) while the statistical power of course decreased. We therefore chose to go with the reviewer-suggested metric of mean transcript position per cell.
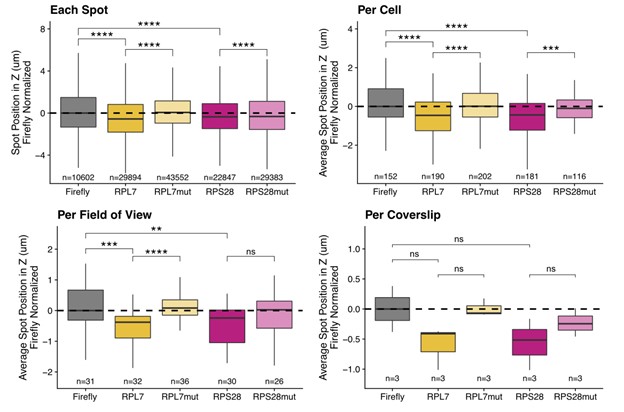
Reviewer Figure 3. C2bbe1 monolayer smFISH spot position analysis. RNA localization across the apicobasal axis is measured by smFISH spot position in the Z axis. This can be plotted for each spot, where thousands of spots over-power the statistics. Spot position can be averaged per cell as outlined manually within the FISH-quant software. This reduces sample size and allows for more accurate statistical analysis. When spot position is averaged per field of view, sample size further decreases, statistics are less powered but the localization trends are still robust. Finally, we can average spot position per coverslip, which represents biological replicates. We lose almost all statistical power as sample size is limited to 3 coverslips. Despite this, the localization trends are still recognizable.
When we use this metric, all results remain the same with the exception of the smFISH validation of endogenous OSBPL3 localization. That result loses its statistical significance and has now been omitted from the manuscript. All epithelial smFISH panels have been updated to use this new metric, and the number of cells associated with each observation is indicated for each sample.
For the neuronal images, these were already quantified at the per-cell level as we compare soma and neurite transcript counts from the same cell. In lieu of more imaging of these samples, we chose to perform subcellular fractionation into soma and neurite samples followed by RT-qPCR as an orthogonal technique (figure 3K, figure 3 supplement 14). This technique profiles the population average of approximately 3 million cells.
- F: How was the segmentation of soma vs. neurites performed? It would be good to have a larger image as a supplemental figure so that it is clear the proximal or distal neurites segments are being compared
All neurite vs. soma segmentations were done manually. An example of this segmentation is included as Reviewer Figure 4. This means that often only proximal neurites segments are included in the analysis as it is often difficult to find an entire soma and an entire neurite in one field of view. However, in our experience, inclusion of more distal neurite segments would likely only strengthen the smFISH results as we often observe many molecules of localized transcripts in the distal tips of these neurites.
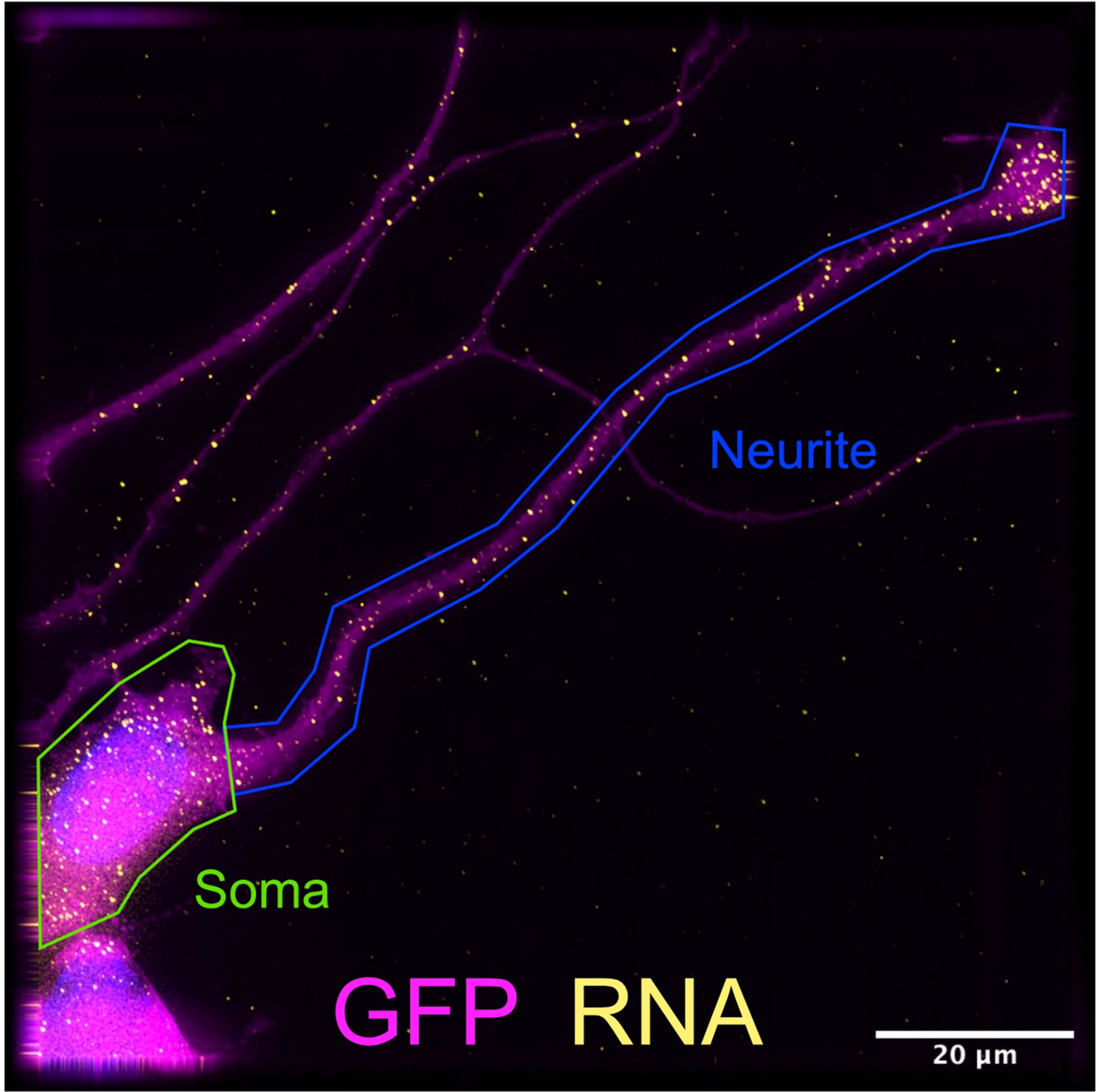
Reviewer Figure 4. Manual segmentation of differentiated CAD soma and neurite in FISH-quant software. Neurites that do not overlap adjacent neurites are selected for imaging. Often neurites extend beyond the field of view, limiting this assay to RNA localization in proximal neurites.
Also, it should be noted that the neuronal smFISH results are now supplemented by experiments involving subcellular fractionation and RT-qPCR (figure 3 supplement 14). These subcellular fractionation experiments collect the whole neurite, both the proximal and distal portions.
Text has been added to the methods under the header “smFISH computational analysis” to clarify how the segmentation was done.
-
Evaluation Summary:
Goering and colleagues investigate subcellular RNA localization across different cell types and species. The major insight is that there may be general mechanisms and specifically conserved proteins that regulate RNA localization in diverse cell types and morphologies. This manuscript will be of interest to those studying gene expression and how its regulation occurs within the cell.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
In this manuscript, Goering et al. investigate subcellular RNA localization across different cell types focusing on epithelial cells (mouse C2bbe1 and human HCA-7 enterocyte monolayers, canine MDCK epithelial cells) as well as neuronal cultures (mouse CAD cells). They use their recently established Halo-seq method to investigate transcriptome-wide RNA localization biases in C2bbe1 enterocyte monolayers and find that 5'TOP-motif containing mRNAs, which encode ribosomal proteins (RPs), are enriched on the basal side of these cells. These results are supported by smFISH against endogenous RP-encoding mRNAs (RPL7 and RPS28) as well as Firefly luciferase reporter transcripts with and without mutated 5'TOP sequences. Furthermore, they find that 5'TOP-motifs are not only driving localization to the basal side of …
Reviewer #1 (Public Review):
In this manuscript, Goering et al. investigate subcellular RNA localization across different cell types focusing on epithelial cells (mouse C2bbe1 and human HCA-7 enterocyte monolayers, canine MDCK epithelial cells) as well as neuronal cultures (mouse CAD cells). They use their recently established Halo-seq method to investigate transcriptome-wide RNA localization biases in C2bbe1 enterocyte monolayers and find that 5'TOP-motif containing mRNAs, which encode ribosomal proteins (RPs), are enriched on the basal side of these cells. These results are supported by smFISH against endogenous RP-encoding mRNAs (RPL7 and RPS28) as well as Firefly luciferase reporter transcripts with and without mutated 5'TOP sequences. Furthermore, they find that 5'TOP-motifs are not only driving localization to the basal side of epithelial cells but also to neuronal processes. To investigate the molecular mechanism behind the observed RNA localization biases, they reduce expression of several Larp proteins and find that RNA localization is consistently Larp1-dependent. Additionally, the localization depends on the placement of the TOP sequence in the 5'UTR and not the 3'UTR. To confirm that similar RNA localization biases can be conserved across cell types for other classes of transcripts, they perform similar experiments with a GA-rich element containing Net1 3'UTR transcript, which has previously been shown to exhibit a strong localization bias in several cell types. In order to determine if motor proteins contribute to these RNA distributions, they use motor protein inhibitors to confirm that the localization of individual members of both classes of transcripts, 5'TOP and GA-rich, is kinesin-dependent and that RNA localization to specific subcellular regions is likely to coincide with RNA localization to microtubule plus ends that concentrate in the basal side of epithelial cells as well as in neuronal processes.
In summary, Goering et al. present an interesting study that contributes to our understanding of RNA localization. While RNA localization has predominantly been studied in a single cell type or experimental system, this work looks for commonalities to explain general principles. I believe that this is an important advance, but there are several points that should be addressed.
Comments:
1. The Mili lab has previously characterized the localization of ribosomal proteins and NET1 to protrusions (Wang et al, 2017, Moissoglu et al 2019, Crisafis et al., 2020) and the role of kinesins in this localization (Pichon et al, 2021). These papers should be cited and their work discussed. I do not believe this reduces the novelty of this study and supports the generality of the RNA localization patterns to additional cellular locations in other cell types.
2. The 5'TOP motif begins with an invariant C nucleotide and mutation of this first nucleotide next to the cap has been shown to reduce translation regulation during mTOR inhibition (Avni et al, 1994 and Biberman et al 1997) and also Lapr1 binding (Lahr et al, 2017). Consequently, it is not clear to me if RPS28 initiates transcription with an A as indicated in Figure 3B. There also seems to be some differences in published CAGE datasets, but this point needs to be clarified. Additionally, it is not clear to me how the 5'TOP Firefly luciferase reporters were generated and if the transcription start site and exact 5'-ends of these constructs were determined. This is again essential to determine if it is a pyrimidine sequence in the 5'UTR that is important for localization or the 5'TOP motif and if Larp1 is directly regulating the localization by binding to the 5'TOP motif or if the effect they observe is indirect (e.g. is Larp1 also basally localized?). It should also be noted that Larp1 has been suggested to bind pyrimidine-rich sequences in the 5'UTR that are not next to the cap, but the details of this interaction are less clear (Al-Ashtal et al, 2021)
3. In figure 1A, they indicate that mRNA stability can contribute to RNA localization, but this point is never discussed. This may be important to their work since Larp1 has also been found to impact mRNA half-lives (Aoki et al, 2013 and Mattijssen et al 2020, Al-Ashtal et al 2021). Is it possible the effect they see when Larp1 is depleted comes from decreased stability?
4. Also Moor et al, 2017 saw that feeding cycles changed the localization of 5'TOP mRNAs. Similarly, does mTOR inhibition or activation or simply active translation alter the localization patterns they observe? Further evidence for dynamic regulation of RNA localization would strengthen this paper
5. For smFISH quantification, is every mRNA treated as an independent measurement so that the statistics are calculated on hundreds of mRNAs? Large sample sizes can give significant p-values but have very small differences as observe for Firefly vs. OSBPL3 localization. Since determining the biological interpretation of effect size is not always clear, I would suggest plotting RNA position per cell or only treat biological replicates as independent measurements to determine statistical significance. This should also be done for other smFISH comparisons
6. F: How was the segmentation of soma vs. neurites performed? It would be good to have a larger image as a supplemental figure so that it is clear the proximal or distal neurites segments are being compared
-
Reviewer #2 (Public Review):
In this work, Goering and colleagues investigated three fundamental aspects of mRNA localization:
First, whether RNA elements previously shown to control subcellular localization of mRNAs in mammalian cells (i.e. TOP, GA, and NET1 3'UTR), are capable of promoting subcellular RNA localization across species (i.e. mouse or human cell lines) and cell types (i.e. epithelial and neuronal cells).
Second, whether RNA binding proteins (RBPs) belonging to the conserved La superfamily, play a role in RNA localization across species and cell types.
Third, whether specific motor protein classes (i.e. kinesins and dyneins) regulate the asymmetric localization of RNAs across species and cell types.By using a combination of RNA-seq-based approaches and reporter mRNAs coupled to RNA imaging, the authors show that mRNAs …
Reviewer #2 (Public Review):
In this work, Goering and colleagues investigated three fundamental aspects of mRNA localization:
First, whether RNA elements previously shown to control subcellular localization of mRNAs in mammalian cells (i.e. TOP, GA, and NET1 3'UTR), are capable of promoting subcellular RNA localization across species (i.e. mouse or human cell lines) and cell types (i.e. epithelial and neuronal cells).
Second, whether RNA binding proteins (RBPs) belonging to the conserved La superfamily, play a role in RNA localization across species and cell types.
Third, whether specific motor protein classes (i.e. kinesins and dyneins) regulate the asymmetric localization of RNAs across species and cell types.By using a combination of RNA-seq-based approaches and reporter mRNAs coupled to RNA imaging, the authors show that mRNAs with specific elements (TOP, GA, and NET1 3'UTR) localize asymmetrically across species and cell types and that this is likely mediated by conserved RBPs and via direct transport mechanisms preferentially involving kinesin motors.
Major strengths of this work:
- The systematic comparison across species and cell types of the distribution of mRNA localization reporters and the role of specific RBPs and motor proteins during RNA localization.
- The combination of genome-wide (i.e. HALO-seq) and RNA imaging approaches (smFISH, smiFISH).
- The results reported in this study represent a significant conceptual advancement.Major weaknesses of this work:
- The systematic comparison or RNA localization across species and cell types done by RNA imaging is performed only using RNA localization reporters and not on endogenous genes. Thus, a validation on endogenous transcripts is missing.
- The systematic characterization of the reporter RNA localization strongly relies on RNA imaging approaches (smFISH, smiFISH). However, the number of cells counted for each experiment is very limited (usually ~10 cells) and it is not clear if the quantifications derive from independent experiments.Overall, the manuscript is well written and clear. The data presented in this work support the claims made by the authors, even though the mRNA imaging data should be strengthened by additional quantifications.
The impact of this work on the RNA localization field is two-fold; First, the RNA-seq-based data and the bioinformatics analysis, comparing localized RNAs across cell types and species, provide a rich list of RNAs that could be investigated in future studies.
Second, the results of this study provide a substantial conceptual novelty: that is that for specific mRNA classes, such as ribosomal protein-encoding mRNAs, the mechanisms controlling RNA localization and transport are highly conserved suggesting that they represent early evolved mechanisms to regulate gene expression.
As noted by the authors, it is important to highlight that ribosomes are assembled in the nucleus, thus it remains very unclear the reason why mammalian cells show such strong conservation in the localization far from the nucleus (e.g. in neurites) of mRNAs encoding ribosomal protein genes. Future studies should elucidate the role of this localization and whether it promotes ribosome heterogeneity and function. -
