Representational integration and differentiation in the human hippocampus following goal-directed navigation
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
This is a carefully designed and analysed fMRI study investigating how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. It will be of much interest to researchers studying the differentiation and integration of memories and the formation of cognitive maps. The results provide new insight into how local and global knowledge about our environment is represented, but some of the conclusions and interpretations could be strengthened with additional analyses.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 and Reviewer #3 agreed to share their names with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
As we learn, dynamic memory processes build structured knowledge across our experiences. Such knowledge enables the formation of internal models of the world that we use to plan, make decisions, and act. Recent theorizing posits that mnemonic mechanisms of differentiation and integration – which at one level may seem to be at odds – both contribute to the emergence of structured knowledge. We tested this possibility using fMRI as human participants learned to navigate within local and global virtual environments over the course of 3 days. Pattern similarity analyses on entorhinal cortical and hippocampal patterns revealed evidence that differentiation and integration work concurrently to build local and global environmental representations, and that variability in integration relates to differences in navigation efficiency. These results offer new insights into the neural machinery and the underlying mechanisms that translate experiences into structured knowledge that allows us to navigate to achieve goals.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
This is a carefully-conducted fMRI study looking at how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. Collectively, the results from the study provide valuable additional constraints on our understanding of representational change in the medial temporal lobes and spatial learning. The most notable finding is that representational similarity in the hippocampus post-local-learning (but prior to any global navigation trials) predicts the efficiency of subsequent global navigation.
Strengths:
The paper has several strengths. It uses a clever two-phase paradigm that makes it possible to track how participants learn local structure as well as how they piece together global structure …
Author Response
Reviewer #1 (Public Review):
This is a carefully-conducted fMRI study looking at how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. Collectively, the results from the study provide valuable additional constraints on our understanding of representational change in the medial temporal lobes and spatial learning. The most notable finding is that representational similarity in the hippocampus post-local-learning (but prior to any global navigation trials) predicts the efficiency of subsequent global navigation.
Strengths:
The paper has several strengths. It uses a clever two-phase paradigm that makes it possible to track how participants learn local structure as well as how they piece together global structure based on exposure to local environments. Using this paradigm, the authors show that - after local learning - hippocampal representations of landmarks that appeared within the same local environment show differentiation (i.e., neural similarity is higher for more distant landmarks) but landmarks that appeared in different local environments show the opposite pattern of results (i.e., neural similarity is lower for more distant landmarks); after participants have the opportunity to navigate globally, the latter finding goes away (i.e., neural similarity for landmarks that occurred in different local environments is no longer influenced by the distance between landmarks). Lastly, the authors show that the degree of hippocampal sensitivity to global distance after local-only learning (but before participants have the opportunity to navigate globally) negatively predicts subsequent global navigation efficiency. Taken together, these results meaningfully extend the space of data that can be used to constrain theories of MTL contributions to spatial learning.
We appreciate Dr. Norman’s generous feedback here along with his other insightful comments. Please see below for a point-by-point response. We note that responses to a number of Dr. Norman’s points were surfaced by the Editor as Essential revisions; as such, in a number of instances in the point-by-point below we direct Dr. Norman to our responses above under the Essential revisions section.
Weaknesses:
General comment 1: The study has an exploratory feel, in the sense that - for the most part - the authors do not set forth specific predictions or hypotheses regarding the results they expected to obtain. When hypotheses are listed, they are phrased in a general way (e.g., "We hypothesized that we would find evidence for both integration and differentiation emerging at the same time points across learning, as participants build local and global representations of the virtual environment", and "We hypothesized that there would be a change in EC and hippocampal pattern similarity for items located on the same track vs. items located on different tracks" - this does not specify what the change will be and whether the change is expected to be different for EC vs. hippocampus). I should emphasize that this is not, unto itself, a weakness of the study, and it appears that the authors have corrected for multiple comparisons (encompassing the range of outcomes explored) throughout the paper. However, at times it was unclear what "denominator" was being used for the multiple comparisons corrections (i.e., what was the full space of analysis options that was being corrected for) - it would be helpful if the authors could specify this more concretely, throughout the paper.
We appreciate this guidance and the importance of these points. We have taken a number of steps to clarify our hypotheses, we now distinguish a priori predictions from exploratory analyses, and we now explicitly indicate throughout the manuscript how we corrected for multiple comparisons. For full details, please see above for our response to Essential Revisions General comment #1.
General comment 2: Some of the analyses featured prominently in the paper (e.g., interactions between context and scan in EC) did not pass multiple comparisons correction. I think it's fine to include these results in the paper, but it should be made clear whenever they are mentioned that the results were not significant after multiple comparisons correction (e.g., in the discussion, the authors say "learning restructures representations in the hippocampus and in the EC", but in that sentence, they don't mention that the EC results fail to pass multiple comparisons correction).
Thank you for encouraging greater clarity here. As noted directly above, we now explicitly indicate our a priori predictions, we state explicitly which results survive multiple comparisons correction, and we added necessary caveats for effects that should be interpreted with caution.
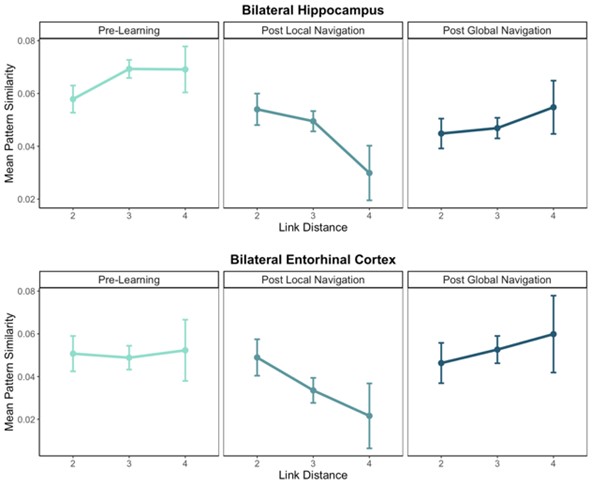
General comment 3: The authors describe the "flat" pattern across the distance 2, 3, and 4 conditions in Figure 4c (post-global navigation) and in Figure 5b (in the "more efficient" group) as indicating integration. However, this flat pattern across 2, 3, and 4 (unto itself) could simply indicate that the region is insensitive to location - is there some other evidence that the authors could bring to bear on the claim that this truly reflects integration? Relatedly, in the discussion, the authors say "the data suggest that, prior to Global Navigation, LEs had integrated only the nearest landmarks located on different tracks (link distance 2)" - what is the basis for this claim? Considered on its own, the fact that similarity was high for link distance 2 does not indicate that integration took place. If the authors cannot get more direct evidence for integration, it might be useful for them to hedge a bit more in how they interpret the results (the finding is still very interesting, regardless of its cause).
Based on the outcomes of additional behavioral and neural analyses that were helpfully suggested by reviewers, we revised discussion of this aspect of the data. Please see our response above under Essential Revisions General comment #4 for full details of the changes made to the manuscript.
Reviewer #2 (Public Review):
This paper presents evidence of neural pattern differentiation (using representational similarity analysis) following extensive experience navigating in virtual reality, building up from individual tracks to an overall environment. The question of how neural patterns are reorganized following novel experiences and learning to integrate across them is a timely and interesting one. The task is carefully designed and the analytic setup is well-motivated. The experimental approach provides a characterization of the development of neural representations with learning across time. The behavioral analyses provide helpful insight into the participants' learning. However, there were some aspects of the conceptual setup and the analyses that I found somewhat difficult to follow. It would also be helpful to provide clearer links between specific predictions and theories of hippocampal function.
We appreciate the Reviewer’s careful read of our manuscript and their thoughtful guidance for improvement, which we believe strengthened the revised product. We note that responses to a number of the Reviewer’s points were surfaced by the Editor as Essential revisions; as such, in a number of instances in the point-by-point below we direct the Reviewer to our responses above under the Essential revisions section.
General comment 1: The motivation in the Introduction builds on the assumption that global representations are dependent on local ones. However, I was not completely sure about the specific predictions or assumptions regarding integration vs. differentiation and their time course in the present experimental design. What would pattern similarity consistent with 'early evidence of global map learning' (p. 7) look like? Fig. 1D was somewhat difficult to understand. The 'state space' representation is only shown in Figure 1 while all subsequent analyses are averaged pairwise correlations. It would be helpful to spell out predictions as they relate to the similarity between same-route vs. different-route neural patterns.
We appreciate this feedback. An increase in pattern similarity across features that span tracks would indicate the linking of those features together. ‘Early evidence’ here describes the point in experience where participants had traversed local (within-track) paths but had yet to traverse across-tracks.
Figure 1D seeks to communicate the high-level conceptual point about how similarity (abstractly represented as state-space distance) may change in one of two directions as a function of experience.
General comment 2: The shared landmarks could be used by the participants to infer how the three tracks connected even before they were able to cross between them. It is possible that the more efficient navigators used an explicit encoding strategy to help them build a global map of the world. While I understand the authors' reasoning for excluding the shared landmarks (p. 13), it seems like it could be useful to run an analysis including them as well - one possibility is that they act as 'anchors' and drive the similarity between different tracks early on; another is that they act as 'boundaries' and repel the representations across routes. Assuming that participants crossed over at these landmarks, these seem like particularly salient aspects of the environment.
We agree that these shared landmarks play an important role in learning the global environment and guiding participants’ navigation. However, they also add confounding elements to the analyses; mainly, shared landmarks are located near multiple goal locations and associated with multiple tracks, and transition probabilities differ at shared landmarks because they have an increased number of neighboring landmarks and fractals. In the initial submission, shared landmarks were included in all analyses except (a) global distance models and (b) context models (which compare items located on the same vs different tracks).
With respect to (a) the global distance models, we ran these models while including shared landmarks and the results did not differ (see figure below and compare to Fig. 5 in the revised manuscript):
Distance representations in the Global Environment, with shared landmarks included. These data can be compared to Figure 5 of the revised manuscript, which does not include shared landmarks (see page 5 of this response letter).
We continue to report the results from models excluding shared landmarks due to the confounding factors described above, with the following addition to the Results section:
“We excluded shared landmarks from this model as they are common to multiple tracks; however, the results do not differ if these landmarks are included in the analysis.”
With respect to (b) the context analyses (which compare items located on the same vs different tracks), we cannot include shared landmarks in these analyses because they are common amongst multiple tracks and thus confound the analyses. Finally, we are unable to conduct additional analyses investigating shared landmarks specifically (for example, examining how similarity between shared landmarks evolves across learning) due to very low trial counts. We share the Reviewer’s perspective that the role of shared landmarks during the building of map representations promises to provide additional insights and believe this is a promising question for future investigation.
General comment 3: What were the predictions regarding the fractals vs. landmarks (p. 13)? It makes sense to compare like-to-like, but since both were included in the models it would be helpful to provide predictions regarding their similarity patterns.
We are grateful for the feedback on how to improve the consistency of results reporting. In the revision, we updated the relevant sections of the manuscript to include results from fractals. Please see our above response to Essential Revisions General comment #5 for additions made to the text.
General comment 4: The median split into less-efficient and more-efficient groups does not seem to be anticipated in the Introduction and results in a small-N group comparison. Instead, as the authors have a wealth of within-individual data, it might be helpful to model single-trial navigation data in relation to pairwise similarity values for each given pair of landmarks in a mixed-effects model. While there won't be a simple one-to-one mapping and fMRI data are noisy, this approach would afford higher statistical power due to more within-individual observations and would avoid splitting the sample into small subgroups.
We appreciate this very helpful suggestion. Following this guidance, we removed the median-split analysis and ran a mixed-effects model relating trial-wise navigation data (at the beginning of the Global Navigation Task) to pairwise similarity values for each given pair of landmarks and fractals (Post Local Navigation). We also altered our approach to the across-participant analysis examining brain-behavior relationships. Please see our above response to Essential Revisions General comment #3 for additions to the revised manuscript.
General comment 5: If I understood correctly, comparing Fig. 4B and Fig. 5B suggests that the relationship between higher link distance and lower representational similarity was driven by less efficient navigators. The performance on average improved over time to more or less the same level as within-track (Fig. 2). Were less efficient navigators particularly inefficient on trials with longer distances? In the context of models of hippocampal function, this suggests that good navigators represented all locations as equidistant while poorer navigators showed representations more consistent with a map - locations that were further apart were more distant in their representational patterns. Perhaps more fine-grained analyses linking neural patterns to behavior would be helpful here.
Following the above guidance, we removed the median-split analyses when exploring across-participant brain-behavior relationships (see Essential Revisions General comment #3), replacing it with a mixed-effects model analysis, and we revised our discussion of the across-track link distance effects (see Essential Revisions General comment #4). For this reason, we were hesitant and ultimately decided against conducting the proposed fine-grained analyses on the median-split data.
General comment 6: I'm not completely sure how to interpret the functional connectivity analysis between the vmPFC and the hippocampus vs. visual cortex (Fig. 6). The analysis shows that the hippocampus and visual cortex are generally more connected than the vmPFC and visual cortex - but this relationship does not show an experience-dependent relationship and is consistent with resting-state data where the hippocampus tends to cluster into the posterior DMN network.
We expected to see an experience-dependent relationship between vmPFC and hippocampal pattern similarity, and agree that these findings are difficult to interpret. Based on comments from several reviewers, we removed the second-order similarity analysis from the manuscript in favor of an analysis which models the relationship between vmPFC pattern similarity and hippocampal pattern similarity. Moreover, given the exploratory nature of the vmPFC analyses, and following guidance from Reviewer 1 about the visual cortex control analyses, both were moved to the Appendix. Please see our above response to Essential Revisions General comment #7 for further details of the changes made to the manuscript.
Reviewer #3 (Public Review):
Fernandez et al. report results from a multi-day fMRI experiment in which participants learned to locate fractal stimuli along three oval-shaped tracks. The results suggest the concurrent emergence of a local, differentiated within-track representation and a global, integrated cross-track representation. More specifically, the authors report decreases in pattern similarity for stimuli encountered on the same track in the entorhinal cortex and hippocampus relative to a pre-task baseline scan. Intriguingly, following navigation on the individual tracks, but prior to global navigation requiring track-switching, pattern similarity in the hippocampus correlated with link distances between landmark stimuli. This effect was only observed in participants who navigated less efficiently in the global navigation task and was absent after global navigation.
Overall, the study is of high quality in my view and addresses relevant questions regarding the differentiation and integration of memories and the formation of so-called cognitive maps. The results reported by the authors are interesting and are based upon a well-designed experiment and thorough data analysis using appropriate techniques. A more detailed assessment of strengths and weaknesses can be found below.
Strengths
- The authors address an interesting question at the intersection of memory differentiation and integration. The study is further relevant for researchers interested in the question of how we form cognitive maps of space.
- The study is well-designed. In particular, the pre-learning baseline scan and the random-order presentation of stimuli during MR scanning allow the authors to track the emergence of representations in a well-controlled fashion. Further, the authors include an adequate control region and report direct comparisons of their effects against the patterns observed in this control region.
- The manuscript is well-written. The introduction provides a good overview of the research field and the discussion does a good job of summarizing the findings of the present study and positioning them in the literature.
We thank Dr. Bellmund for his positive evaluation of the manuscript. We greatly appreciate the insightful feedback, which we believe strengthened the manuscript’s clarity and potential impact. We note that responses to a number of Dr. Bellmund’s points were surfaced by the Editor as Essential revisions; as such, in a number of instances in the point-by-point below we direct the Reviewer to our responses above under the Essential revisions section.
Weaknesses
General comment 1: Despite these distinct strengths, the present study also has some weaknesses. On the behavioral level, I am wondering about the use of path inefficiency as a metric for global navigation performance. Because it is quantified based on the local response, it conflates the contributions of local and global errors.
We appreciate this point with respect to path inefficiency during global navigation. As noted below, following Dr. Bellmund’s further insightful guidance, we now complement the path inefficiency analyses with additional metrics of across-track (global) navigation performance, which effectively separate local from global errors (please see below response to Author recommendation #1).
General comment 2: For the distance-based analysis in the hippocampus, the authors choose to only analyze landmark images and do not include fractal stimuli. There seems to be little reason to expect that distances between the fractal stimuli, on which the memory task was based, would be represented differently relative to distances between the landmarks.
We are grateful for the feedback on how to improve the consistency of results reporting. In the revision, we updated the relevant sections of the manuscript to include results from fractals. Please see our above response to Essential Revisions General comment #5 for full details.
General comment 3: Related to the aforementioned analysis, I am wondering why the authors chose the link distance between landmarks as their distance metric for the analysis and why they limit their analysis to pairs of stimuli with distance 1 or 2 and do not include pairs separated by the highest possible distance (3).
We appreciate the request for clarification here. Beginning with the latter question, we note that the highest possible distance varies between within-track vs. across-track paths. If participants navigate in the Local Navigation Task using the shortest or most efficient path, the highest possible within-track link distance between two stimuli is 2. For this reason, the Local Navigation/within-track analysis includes link distances of 1 and 2. For the Global Navigation analysis, we also include pairs of stimuli with link distances of 3 and 4 when examining across-track landmarks.
Regarding the use of link distance as the distance metric, we note that the path distance (a.u.) varies only slightly between pairs of stimuli with the same link distance. As such, categorical treatment link distance accounts for the vast majority of the variance in path distance and thus is a suitable approach. Please note that in the new trial-level brain-behavior analysis included in the revised manuscript (which replaces the median-split analysis), we used the length of the optimal path.
General comment 4: Surprisingly, the authors report that across-track distances can be observed in the hippocampus after local navigation, but that this effect cannot be detected after global, cross-track navigation. Relatedly, the cross-track distance effect was detected only in the half of participants that performed relatively badly in the cross-track navigation task. In the results and discussion, the authors suggest that the effect of cross-track distances cannot be detected because participants formed a "more fully integrated global map". I do not find this a convincing explanation for why the effect the authors are testing would be absent after global navigation and for why the effect was only present in those participants who navigated less efficiently.
We appreciate Dr. Bellmund’s input here, which was shared by other reviewers. We revised and clarified the Discussion based on reviewer comments. Please see our above response to Essential Revisions General comment #4 for full details.
General comment 5: The authors report differences in the hippocampal representational similarity between participants who navigated along inefficient vs. efficient paths. These are based on a median split of the sample, resulting in a comparison of groups including 11 and 10 individuals, respectively. The median split (see e.g. MacCallum et al., Psychological Methods, 2002) and the low sample size mandate cautionary interpretation of the resulting findings about interindividual differences.
We appreciate the feedback we received from multiple reviewers with respect to the median-split brain-behavior analysis. We replaced the median-split analysis with the following: 1) a mixed-effects model predicting neural pattern similarity Post Local Navigation, with a continuous metric of task performance (each participant’s median path inefficiency for across-track trials in the first four test runs of Global Navigation) and link distance as predictors; and 2) a mixed-effects model relating trial-wise navigation data to pairwise similarity values for each given pair of landmarks and fractals (as suggested by Reviewer 2). Please see our above response to Essential Revisions General comment #3 for additions to the revised manuscript.
-
Evaluation Summary:
This is a carefully designed and analysed fMRI study investigating how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. It will be of much interest to researchers studying the differentiation and integration of memories and the formation of cognitive maps. The results provide new insight into how local and global knowledge about our environment is represented, but some of the conclusions and interpretations could be strengthened with additional analyses.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 and Reviewer #3 agreed to share their names with the authors.)
-
Reviewer #1 (Public Review):
This is a carefully-conducted fMRI study looking at how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. Collectively, the results from the study provide valuable additional constraints on our understanding of representational change in the medial temporal lobes and spatial learning. The most notable finding is that representational similarity in the hippocampus post-local-learning (but prior to any global navigation trials) predicts the efficiency of subsequent global navigation.
Strengths:
The paper has several strengths. It uses a clever two-phase paradigm that makes it possible to track how participants learn local structure as well as how they piece together global structure based on exposure to …
Reviewer #1 (Public Review):
This is a carefully-conducted fMRI study looking at how neural representations in the hippocampus, entorhinal cortex, and ventromedial prefrontal cortex change as a function of local and global spatial learning. Collectively, the results from the study provide valuable additional constraints on our understanding of representational change in the medial temporal lobes and spatial learning. The most notable finding is that representational similarity in the hippocampus post-local-learning (but prior to any global navigation trials) predicts the efficiency of subsequent global navigation.
Strengths:
The paper has several strengths. It uses a clever two-phase paradigm that makes it possible to track how participants learn local structure as well as how they piece together global structure based on exposure to local environments. Using this paradigm, the authors show that - after local learning - hippocampal representations of landmarks that appeared within the same local environment show differentiation (i.e., neural similarity is higher for more distant landmarks) but landmarks that appeared in different local environments show the opposite pattern of results (i.e., neural similarity is lower for more distant landmarks); after participants have the opportunity to navigate globally, the latter finding goes away (i.e., neural similarity for landmarks that occurred in different local environments is no longer influenced by the distance between landmarks). Lastly, the authors show that the degree of hippocampal sensitivity to global distance after local-only learning (but before participants have the opportunity to navigate globally) negatively predicts subsequent global navigation efficiency. Taken together, these results meaningfully extend the space of data that can be used to constrain theories of MTL contributions to spatial learning.
Weaknesses:
1. The study has an exploratory feel, in the sense that - for the most part - the authors do not set forth specific predictions or hypotheses regarding the results they expected to obtain. When hypotheses are listed, they are phrased in a general way (e.g., "We hypothesized that we would find evidence for both integration and differentiation emerging at the same time points across learning, as participants build local and global representations of the virtual environment", and "We hypothesized that there would be a change in EC and hippocampal pattern similarity for items located on the same track vs. items located on different tracks" - this does not specify what the change will be and whether the change is expected to be different for EC vs. hippocampus). I should emphasize that this is not, unto itself, a weakness of the study, and it appears that the authors have corrected for multiple comparisons (encompassing the range of outcomes explored) throughout the paper. However, at times it was unclear what "denominator" was being used for the multiple comparisons corrections (i.e., what was the full space of analysis options that was being corrected for) - it would be helpful if the authors could specify this more concretely, throughout the paper.
2. Some of the analyses featured prominently in the paper (e.g., interactions between context and scan in EC) did not pass multiple comparisons correction. I think it's fine to include these results in the paper, but it should be made clear whenever they are mentioned that the results were not significant after multiple comparisons correction (e.g., in the discussion, the authors say "learning restructures representations in the hippocampus and in the EC", but in that sentence, they don't mention that the EC results fail to pass multiple comparisons correction).
3. The authors describe the "flat" pattern across the distance 2, 3, and 4 conditions in Figure 4c (post-global navigation) and in Figure 5b (in the "more efficient" group) as indicating integration. However, this flat pattern across 2, 3, and 4 (unto itself) could simply indicate that the region is insensitive to location - is there some other evidence that the authors could bring to bear on the claim that this truly reflects integration? Relatedly, in the discussion, the authors say "the data suggest that, prior to Global Navigation, LEs had integrated only the nearest landmarks located on different tracks (link distance 2)" - what is the basis for this claim? Considered on its own, the fact that similarity was high for link distance 2 does not indicate that integration took place. If the authors cannot get more direct evidence for integration, it might be useful for them to hedge a bit more in how they interpret the results (the finding is still very interesting, regardless of its cause).
-
Reviewer #2 (Public Review):
This paper presents evidence of neural pattern differentiation (using representational similarity analysis) following extensive experience navigating in virtual reality, building up from individual tracks to an overall environment. The question of how neural patterns are reorganized following novel experiences and learning to integrate across them is a timely and interesting one. The task is carefully designed and the analytic setup is well-motivated. The experimental approach provides a characterization of the development of neural representations with learning across time. The behavioral analyses provide helpful insight into the participants' learning. However, there were some aspects of the conceptual setup and the analyses that I found somewhat difficult to follow. It would also be helpful to provide …
Reviewer #2 (Public Review):
This paper presents evidence of neural pattern differentiation (using representational similarity analysis) following extensive experience navigating in virtual reality, building up from individual tracks to an overall environment. The question of how neural patterns are reorganized following novel experiences and learning to integrate across them is a timely and interesting one. The task is carefully designed and the analytic setup is well-motivated. The experimental approach provides a characterization of the development of neural representations with learning across time. The behavioral analyses provide helpful insight into the participants' learning. However, there were some aspects of the conceptual setup and the analyses that I found somewhat difficult to follow. It would also be helpful to provide clearer links between specific predictions and theories of hippocampal function.
1. The motivation in the Introduction builds on the assumption that global representations are dependent on local ones. However, I was not completely sure about the specific predictions or assumptions regarding integration vs. differentiation and their time course in the present experimental design. What would pattern similarity consistent with 'early evidence of global map learning' (p. 7) look like? Fig. 1D was somewhat difficult to understand. The 'state space' representation is only shown in Figure 1 while all subsequent analyses are averaged pairwise correlations. It would be helpful to spell out predictions as they relate to the similarity between same-route vs. different-route neural patterns.
2. The shared landmarks could be used by the participants to infer how the three tracks connected even before they were able to cross between them. It is possible that the more efficient navigators used an explicit encoding strategy to help them build a global map of the world. While I understand the authors' reasoning for excluding the shared landmarks (p. 13), it seems like it could be useful to run an analysis including them as well - one possibility is that they act as 'anchors' and drive the similarity between different tracks early on; another is that they act as 'boundaries' and repel the representations across routes. Assuming that participants crossed over at these landmarks, these seem like particularly salient aspects of the environment.
3. What were the predictions regarding the fractals vs. landmarks (p. 13)? It makes sense to compare like-to-like, but since both were included in the models it would be helpful to provide predictions regarding their similarity patterns.
4. The median split into less-efficient and more-efficient groups does not seem to be anticipated in the Introduction and results in a small-N group comparison. Instead, as the authors have a wealth of within-individual data, it might be helpful to model single-trial navigation data in relation to pairwise similarity values for each given pair of landmarks in a mixed-effects model. While there won't be a simple one-to-one mapping and fMRI data are noisy, this approach would afford higher statistical power due to more within-individual observations and would avoid splitting the sample into small subgroups.
5. If I understood correctly, comparing Fig. 4B and Fig. 5B suggests that the relationship between higher link distance and lower representational similarity was driven by less efficient navigators. The performance on average improved over time to more or less the same level as within-track (Fig. 2). Were less efficient navigators particularly inefficient on trials with longer distances? In the context of models of hippocampal function, this suggests that good navigators represented all locations as equidistant while poorer navigators showed representations more consistent with a map - locations that were further apart were more distant in their representational patterns. Perhaps more fine-grained analyses linking neural patterns to behavior would be helpful here.
6. I'm not completely sure how to interpret the functional connectivity analysis between the vmPFC and the hippocampus vs. visual cortex (Fig. 6). The analysis shows that the hippocampus and visual cortex are generally more connected than the vmPFC and visual cortex - but this relationship does not show an experience-dependent relationship and is consistent with resting-state data where the hippocampus tends to cluster into the posterior DMN network.
-
Reviewer #3 (Public Review):
Fernandez et al. report results from a multi-day fMRI experiment in which participants learned to locate fractal stimuli along three oval-shaped tracks. The results suggest the concurrent emergence of a local, differentiated within-track representation and a global, integrated cross-track representation. More specifically, the authors report decreases in pattern similarity for stimuli encountered on the same track in the entorhinal cortex and hippocampus relative to a pre-task baseline scan. Intriguingly, following navigation on the individual tracks, but prior to global navigation requiring track-switching, pattern similarity in the hippocampus correlated with link distances between landmark stimuli. This effect was only observed in participants who navigated less efficiently in the global navigation task …
Reviewer #3 (Public Review):
Fernandez et al. report results from a multi-day fMRI experiment in which participants learned to locate fractal stimuli along three oval-shaped tracks. The results suggest the concurrent emergence of a local, differentiated within-track representation and a global, integrated cross-track representation. More specifically, the authors report decreases in pattern similarity for stimuli encountered on the same track in the entorhinal cortex and hippocampus relative to a pre-task baseline scan. Intriguingly, following navigation on the individual tracks, but prior to global navigation requiring track-switching, pattern similarity in the hippocampus correlated with link distances between landmark stimuli. This effect was only observed in participants who navigated less efficiently in the global navigation task and was absent after global navigation.
Overall, the study is of high quality in my view and addresses relevant questions regarding the differentiation and integration of memories and the formation of so-called cognitive maps. The results reported by the authors are interesting and are based upon a well-designed experiment and thorough data analysis using appropriate techniques. A more detailed assessment of strengths and weaknesses can be found below.
Strengths
1. The authors address an interesting question at the intersection of memory differentiation and integration. The study is further relevant for researchers interested in the question of how we form cognitive maps of space.
2. The study is well-designed. In particular, the pre-learning baseline scan and the random-order presentation of stimuli during MR scanning allow the authors to track the emergence of representations in a well-controlled fashion. Further, the authors include an adequate control region and report direct comparisons of their effects against the patterns observed in this control region.
3. The manuscript is well-written. The introduction provides a good overview of the research field and the discussion does a good job of summarizing the findings of the present study and positioning them in the literature.
Weaknesses
1. Despite these distinct strengths, the present study also has some weaknesses. On the behavioral level, I am wondering about the use of path inefficiency as a metric for global navigation performance. Because it is quantified based on the local response, it conflates the contributions of local and global errors.
2. For the distance-based analysis in the hippocampus, the authors choose to only analyze landmark images and do not include fractal stimuli. There seems to be little reason to expect that distances between the fractal stimuli, on which the memory task was based, would be represented differently relative to distances between the landmarks.
3. Related to the aforementioned analysis, I am wondering why the authors chose the link distance between landmarks as their distance metric for the analysis and why they limit their analysis to pairs of stimuli with distance 1 or 2 and do not include pairs separated by the highest possible distance (3).
4. Surprisingly, the authors report that across-track distances can be observed in the hippocampus after local navigation, but that this effect cannot be detected after global, cross-track navigation. Relatedly, the cross-track distance effect was detected only in the half of participants that performed relatively badly in the cross-track navigation task. In the results and discussion, the authors suggest that the effect of cross-track distances cannot be detected because participants formed a "more fully integrated global map". I do not find this a convincing explanation for why the effect the authors are testing would be absent after global navigation and for why the effect was only present in those participants who navigated less efficiently.
5. The authors report differences in the hippocampal representational similarity between participants who navigated along inefficient vs. efficient paths. These are based on a median split of the sample, resulting in a comparison of groups including 11 and 10 individuals, respectively. The median split (see e.g. MacCallum et al., Psychological Methods, 2002) and the low sample size mandate cautionary interpretation of the resulting findings about interindividual differences.
-
-
-
-
-
