Monkeys exhibit human-like gaze biases in economic decisions
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
Lupkin and McGinty studied gaze patterns in monkeys during value-based decision-making to determine whether relationships between gaze and choices that have been described in humans are also present in monkeys. Although previous literature has interpreted the effects of gaze on choice behavior to mean that attention to an option influences the decision that is made, a full understanding of the mechanisms underlying these biases will ultimately require data from non-human primates, making this an important and timely study. The authors use a clever task design to show that, as in humans, value-based choices are related to the amount of time spent viewing each option. The study is well-designed and the topic is significant to the field, but reviewers raised concerns about potential confounds and limitations not addressed by the current analyses.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
In economic decision-making individuals choose between items based on their perceived value. For both humans and nonhuman primates, these decisions are often carried out while shifting gaze between the available options. Recent studies in humans suggest that these shifts in gaze actively influence choice, manifesting as a bias in favor of the items that are viewed first, viewed last, or viewed for the overall longest duration in a given trial. This suggests a mechanism that links gaze behavior to the neural computations underlying value-based choices. In order to identify this mechanism, it is first necessary to develop and validate a suitable animal model of this behavior. To this end, we have created a novel value-based choice task for macaque monkeys that captures the essential features of the human paradigms in which gaze biases have been observed. Using this task, we identified gaze biases in the monkeys that were both qualitatively and quantitatively similar to those in humans. In addition, the monkeys’ gaze biases were well-explained using a sequential sampling model framework previously used to describe gaze biases in humans—the first time this framework has been used to assess value-based decision mechanisms in nonhuman primates. Together, these findings suggest a common mechanism that can explain gaze-related choice biases across species, and open the way for mechanistic studies to identify the neural origins of this behavior.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
Point 1: Many of the initial analyses of behavior metrics, for instance predicting reaction times, number of fixations, or fixation duration, use value difference as a regressor. However, given a limited set of values, value differences are highly correlated with the option values themselves, as well as the chosen value. For instance, in this task the only time when there will be a value difference of 4 drops is when the options are 1 and 5 drops, and given the high performance of these monkeys, this means the chosen value will overwhelmingly be 5 drops. Likewise, there are only two combinations that can yield a value difference of 3 (5 vs. 2 and 4 vs 1), and each will have relatively high chosen values. Given that value motivates behavior and attracts attention, it may be that some of the …
Author Response
Reviewer #1 (Public Review):
Point 1: Many of the initial analyses of behavior metrics, for instance predicting reaction times, number of fixations, or fixation duration, use value difference as a regressor. However, given a limited set of values, value differences are highly correlated with the option values themselves, as well as the chosen value. For instance, in this task the only time when there will be a value difference of 4 drops is when the options are 1 and 5 drops, and given the high performance of these monkeys, this means the chosen value will overwhelmingly be 5 drops. Likewise, there are only two combinations that can yield a value difference of 3 (5 vs. 2 and 4 vs 1), and each will have relatively high chosen values. Given that value motivates behavior and attracts attention, it may be that some of the putative effects of choice difficulty are actually driven by value.
To address this question, we have adapted the methods of Balewski and colleagues (Neuron, 2022) to isolate the unique contributions of chosen value and trial difficulty to reaction time and the number of fixations in a given trial (the two behaviors modulated by difficulty in the original paper). This new analysis reveals a double dissociation in which reaction time decreases as a function of chosen value but not difficulty, while the number of fixations in a trial shows the opposite pattern. Our interpretation is that reaction time largely reflects reward anticipation, whereas the number of fixations largely reflects the amount of information required to render a decision (i.e., choice difficulty). See lines 144-167 and Figure 2.
Point 2: Related to point 1, the study found that duration of first fixations increased with fixated values, and second (middle) fixation durations decreased with fixated value but increased with relative value of the fixated versus other value. Can this effect be more concisely described as an effect of the value of the first fixated option carrying over into behavior during the second fixation?
This is a valid interpretation of the results. To test this directly, we now include an analysis of middle fixation duration as a function of the not-currentlyviewed target. Note that the vast majority of middle fixations are the second fixation in the trial, and therefore the value of the unattended target is typically the one that was viewed first. The analysis showed a negative correlation between middle fixation duration and the value of the unattended target which is consistent with the first fixated value carrying over to the second fixation. See lines 243-246.
Point 3: Given that chosen (and therefore anticipated) values can motivate responses, often measured as faster reaction times or more vigorous motor movements, it seems curious that terminal non-decision times were calculated as a single value for all trials. Shouldn't this vary depending at least on chosen values, and perhaps other variables in the trial?
In all sequential sampling model formulations we are aware of, nondecision time is considered to be fixed across trial types. Examples can be found for perceptual decisions (e.g., Resulaj et al., 2009) and in the “bifurcation point” approach used in the recent value-based decision study by Westbrook et al. (2020).
To further investigate this issue, we asked whether other post-decision processes were sensitive to chosen value in our paradigm. To do so, we measured the interval between the center lever lift and the left or right lever press, corresponding to the time taken to perform the reach movement in each trial (reach latency). We then fit a mixed effects model explaining reach latency as a function of chosen value. While the results showed significantly faster reach latencies with higher chosen values, the effect size was very small, showing on average a ~3ms decrease per drop of juice. In other words, between the highest and lowest levels of chosen value (5 vs. 1), there is only a difference of approximately 12ms. In contrast, the main RT measure used in the study (the interval between target onset and center lever lift) is an order of magnitude more sensitive to chosen value, decreasing ~40ms per drop of juice. These results are shown in Author response image 1.
Author response image 1.
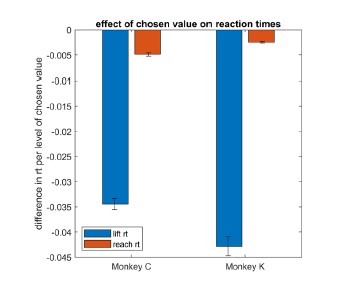
This suggests that post-decision processes (NDT in standard models and the additive stage in the Westbrook paper) vary only minimally as a function of chosen value. We are happy to include this analysis as a supplemental figure upon request.
Point 4: The paper aims to demonstrate similarities between monkey and human gaze behavior in value-based decisions, but focuses mainly on a series of results from one group of collaborators (Krajbich, Rangel and colleagues). Other labs have shown additional nuance that the present data could potentially speak to. First, Cavanaugh et al. (J Exp Psychol Gen, 2014) found that gaze allocation and value differences between options independently influence drift rates on different choices. Second, gaze can correlate with choice because attention to an option amplifies its value (or enhances the accumulation of value evidence) or because chosen options are attended more after the choice is implicitly determined but not yet registered. Westbrook et al. (Science, 2020) found that these effects can be dissociated, with attention influencing choice early in the trial and choice influencing attention later. The NDTs calculated in the present study allot a consistent time to translating a choice into a motor command, but as noted above don't account for potential influences of choice or value on gaze.
The two-stage model of gaze effects put forth by Westbrook et al. (2020) is consistent with other observations of gaze behavior and choice (i.e., Thomas et al., 2019, Smith et al., 2018, Manohar & Husain, 2013). In this model, gaze effects early in the trial are best described by a multiplicative relationship between gaze and value, whereas gaze effects later in the trial are best described with an additive model term. To test the two-stage hypothesis, Westbrook and colleagues determined a ‘bifurcation point’ for each subject that represented the time at which gaze effects transitioned from multiplicative to additive. In our data, trial durations were typically very short (<1s), making it difficult to divide trials and fit separate models to them. We therefore took at different approach: We reasoned that if gaze effects transition from multiplicative to additive at the end of the trial, then the transition point could be estimated by removing data from the end of each trial and assessing the relative fit of a multiplicative vs. additive model. If the early gaze effects are predominantly multiplicative and late gaze effects are additive, the relative goodness of fit for an additive model should decrease as more data are removed from the end of the trial. To test this idea, we compared the relative model fit of an additive vs. multiplicative models in the raw data, and for data in which successively larger epochs were removed from the end of the trial (50, 100, 150, 200, 300, and 400ms). The relative fit was assessed by computing the relative probability that each model accurately reflects the data. In addition, to identify significant differences in goodness of fit, we compared the WAIC values and their standard errors for each model (Supplemental File 3). As shown in Figure 4, the relative fit probability for both models is nonzero in the raw data 0 truncation), indicating that a neither model provides a definitive best fit, potentially reflecting a mixture of the two processes. However, the relative fit of the additive model decreases sharply as data is removed, reaching zero at 100ms truncation. 100ms is also the point at which multiplicative models provide a significantly better fit, indicated by non-overlapping standard error intervals for the two models (Supplemental File 3). Together, this suggested that the transition between early- and late-stage gaze effects likely occurs approximately 100ms before the RT.
To minimize the influence of post-decision gaze effects, the main results use data truncated by 100ms. However, because 100ms is only an estimate, we repeated the main analyses over truncation values between 0 and 400ms, reported in Figure 6 - figure supplement 1 & Figure 7 - figure supplement 1. These show significant gaze duration biases and final gaze biases in data truncated by up to 200ms.
Reviewer #2 (Public Review):
Recommendation 1: The only real issue that I see with the paper is fairly obvious: the authors find that the last fixations are longer than the rest, which is inconsistent with a lot of the human work. They argue that this is due to the reaching required in this task, and they take a somewhat ad-hoc approach to trying to correct for it. Specifically, they take the difference between final and non-final, second fixations, and then choose the 95th percentile of that distribution as the amount of time to subtract from the end of each trial. This amounts to about 200 ms being removed from the end of each trial. There are several issues with this approach. First, it assumes that final and non-final fixations should be the same length, when we know from other work that final fixations are generally shorter. Second, it seems to assume that this 200ms is "the latency between the time that the subject commits to the movement and the time that the movement is actually detected by the experimenter". However, there is a mismatch between that explanation and the details of the task. Those last 200ms are before the monkey releases the middle lever, not before the monkey makes a left/right choice. When the monkey releases the middle lever, the stimuli disappear and they then have 500ms to press the left or right lever. But, the reaction time and fixation data terminate when the monkey releases the middle lever. Consequently, I don't find it very likely that the monkeys are using those last 200ms to plan their hand movement after releasing the middle lever.
Thanks for the opportunity to clarify these points. There are three related issues:
First, with regards to fixation durations, in the updated Figure 3 we now show durations as a function of both the absolute order in the trial (first, second, third, fourth, etc.) and the relative order (final/nonfinal). We find that durations decrease as a function of absolute order in the trial, an effect also seen in humans (see Manohar & Husain, 2013). At the same time, while holding absolute order constant, final fixations are longer than non-final fixations. To explain the discrepancy with human final fixation durations, we note that monkeys make many fewer fixations per trial (~2.5) than humans do (~3.7, computed from publicly available data from Krajbich et al., 2010.) This means that compared to humans, monkeys’ final fixations occur earlier in the trial (e.g., second or third), and are therefore comparatively longer in duration. Note that studies with humans have not independently measured fixation durations by absolute and relative order, and therefore would not have detected the potential interaction between the two effects.
Second, the comment suggests that the final 200ms before lever lift is not spent planning the left/right movement, given that the monkeys have time after the lever lift in which to execute the movement (400 or 500ms, depending on the monkey). The presumption appears to be that 400/500ms should be sufficient to plan a left/right reach. However, we think that these two suggestions are unlikely, and that our original interpretation is the most plausible. First, the 400/500ms deadline between lift and left/right press was set to encourage the monkeys to complete the reach as fast as possible, to minimize deliberations or changes of mind after lifting the lever. More specifically, these deadlines were designed so that on ~0.5% of trials, the monkeys actually fail to complete the reach within the deadline and fail to obtain a reward. This manipulation was effective at motivating fast reaches, as the average reach latency (time between lift and press) was 165 SEM 20ms for Monkey K, and 290 SEM 100ms for Monkey C.
Therefore, given the time pressure imposed by the task, it is very unlikely that significant reach planning occurs after the lever lift. In addition to these empirical considerations, the idea that the final moments before the RT are used for motor planning is a standard assumption in many theoretical models of choice (including sequential sampling models, see Ratcliff & McKoon 2008, for review), and is also well-supported by studies of motor control and motor system neurophysiology. Based on these, we think the assumption of some form of terminal NDT is warranted.
Third, we have changed our method for estimating the NDT interval. In brief we sweep through a range of NDT truncation values (0-400ms) and identify the smallest interval (100ms) that minimizes the contribution of “additive” gaze effects, which are thought to reflect late-stage, post-decision gaze processes. See the response to Point 4 for Reviewer 1 above, Figure 4 and lines 267-325 in the main text. In addition, we report all of the major study results over a range of truncation values between 0 and 400ms.
-
Evaluation Summary:
Lupkin and McGinty studied gaze patterns in monkeys during value-based decision-making to determine whether relationships between gaze and choices that have been described in humans are also present in monkeys. Although previous literature has interpreted the effects of gaze on choice behavior to mean that attention to an option influences the decision that is made, a full understanding of the mechanisms underlying these biases will ultimately require data from non-human primates, making this an important and timely study. The authors use a clever task design to show that, as in humans, value-based choices are related to the amount of time spent viewing each option. The study is well-designed and the topic is significant to the field, but reviewers raised concerns about potential confounds and limitations not …
Evaluation Summary:
Lupkin and McGinty studied gaze patterns in monkeys during value-based decision-making to determine whether relationships between gaze and choices that have been described in humans are also present in monkeys. Although previous literature has interpreted the effects of gaze on choice behavior to mean that attention to an option influences the decision that is made, a full understanding of the mechanisms underlying these biases will ultimately require data from non-human primates, making this an important and timely study. The authors use a clever task design to show that, as in humans, value-based choices are related to the amount of time spent viewing each option. The study is well-designed and the topic is significant to the field, but reviewers raised concerns about potential confounds and limitations not addressed by the current analyses.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
-
Reviewer #1 (Public Review):
McGinty and colleagues studied gaze patterns in monkeys during value-based decision-making. They used a clever task design that obscured items in peripheral vision, encouraging the monkeys to foveate options to collect information about them. They also had monkeys report a decision with their hands to dissociate gaze from choice registration. They report similar relationships between gaze patterns and choices in monkeys as have been previously described in humans, primarily in studies by Krajbich and colleagues, including a bias toward choosing items that were fixated first, last, and longest, which can be captured by variants of a drift diffusion model adapted for gaze-based tasks. Overall, the task design is excellent and the results appear robust, and indeed similar to effects in human studies. However, I …
Reviewer #1 (Public Review):
McGinty and colleagues studied gaze patterns in monkeys during value-based decision-making. They used a clever task design that obscured items in peripheral vision, encouraging the monkeys to foveate options to collect information about them. They also had monkeys report a decision with their hands to dissociate gaze from choice registration. They report similar relationships between gaze patterns and choices in monkeys as have been previously described in humans, primarily in studies by Krajbich and colleagues, including a bias toward choosing items that were fixated first, last, and longest, which can be captured by variants of a drift diffusion model adapted for gaze-based tasks. Overall, the task design is excellent and the results appear robust, and indeed similar to effects in human studies. However, I have some concerns about potential confounds and limitations not currently addressed by the analyses, specifically with respect to the effects of value and choice on gaze metrics.
1. Many of the initial analyses of behavior metrics, for instance predicting reaction times, number of fixations, or fixation duration, use value difference as a regressor. However, given a limited set of values, value differences are highly correlated with the option values themselves, as well as the chosen value. For instance, in this task the only time when there will be a value difference of 4 drops is when the options are 1 and 5 drops, and given the high performance of these monkeys, this means the chosen value will overwhelmingly be 5 drops. Likewise, there are only two combinations that can yield a value difference of 3 (5 vs. 2 and 4 vs 1), and each will have relatively high chosen values. Given that value motivates behavior and attracts attention, it may be that some of the putative effects of choice difficulty are actually driven by value.
2. Related to point 1, the study found that duration of first fixations increased with fixated values, and second (middle) fixation durations decreased with fixated value but increased with relative value of the fixated versus other value. Can this effect be more concisely described as an effect of the value of the first fixated option carrying over into behavior during the second fixation?
3. Given that chosen (and therefore anticipated) values can motivate responses, often measured as faster reaction times or more vigorous motor movements, it seems curious that terminal non-decision times were calculated as a single value for all trials. Shouldn't this vary depending at least on chosen values, and perhaps other variables in the trial?
4. The paper aims to demonstrate similarities between monkey and human gaze behavior in value-based decisions, but focuses mainly on a series of results from one group of collaborators (Krajbich, Rangel and colleagues). Other labs have shown additional nuance that the present data could potentially speak to. First, Cavanaugh et al. (J Exp Psychol Gen, 2014) found that gaze allocation and value differences between options independently influence drift rates on different choices. Second, gaze can correlate with choice because attention to an option amplifies its value (or enhances the accumulation of value evidence) or because chosen options are attended more after the choice is implicitly determined but not yet registered. Westbrook et al. (Science, 2020) found that these effects can be dissociated, with attention influencing choice early in the trial and choice influencing attention later. The NDTs calculated in the present study allot a consistent time to translating a choice into a motor command, but as noted above don't account for potential influences of choice or value on gaze.
-
Reviewer #2 (Public Review):
This article provides evidence that monkeys exhibit human-like gaze biases in value-based decisions. Using a clever experimental design, the authors were able to encourage monkeys to reveal their shifts in attention through eye movements, while they made decisions between juice rewards. Analyzing these eye movements, the authors demonstrate that first fixations, total relative fixation time, and final fixations all predict subsequent choice. One useful feature of their task is that initially the alternatives are masked until the monkey makes a saccade. This means that the first fixations are random with respect to the alternatives, and thus first fixation effects on choice are causal. Moreover, the authors perform an additional manipulation, sometimes staggering the onset of the alternatives, which biases …
Reviewer #2 (Public Review):
This article provides evidence that monkeys exhibit human-like gaze biases in value-based decisions. Using a clever experimental design, the authors were able to encourage monkeys to reveal their shifts in attention through eye movements, while they made decisions between juice rewards. Analyzing these eye movements, the authors demonstrate that first fixations, total relative fixation time, and final fixations all predict subsequent choice. One useful feature of their task is that initially the alternatives are masked until the monkey makes a saccade. This means that the first fixations are random with respect to the alternatives, and thus first fixation effects on choice are causal. Moreover, the authors perform an additional manipulation, sometimes staggering the onset of the alternatives, which biases the first fixation and then the choice.
The only real issue that I see with the paper is fairly obvious: the authors find that the last fixations are longer than the rest, which is inconsistent with a lot of the human work. They argue that this is due to the reaching required in this task, and they take a somewhat ad-hoc approach to trying to correct for it. Specifically, they take the difference between final and non-final, second fixations, and then choose the 95th percentile of that distribution as the amount of time to subtract from the end of each trial. This amounts to about 200 ms being removed from the end of each trial. There are several issues with this approach. First, it assumes that final and non-final fixations should be the same length, when we know from other work that final fixations are generally shorter. Second, it seems to assume that this 200ms is "the latency between the time that the subject commits to the movement and the time that the movement is actually detected by the experimenter". However, there is a mismatch between that explanation and the details of the task. Those last 200ms are before the monkey releases the middle lever, not before the monkey makes a left/right choice. When the monkey releases the middle lever, the stimuli disappear and they then have 500ms to press the left or right lever. But, the reaction time and fixation data terminate when the monkey releases the middle lever. Consequently, I don't find it very likely that the monkeys are using those last 200ms to plan their hand movement after releasing the middle lever.
All that being said, it seems quite unlikely that this particular choice will have any major impact on the qualitative results reported in the paper. This adjustment has no effect on the initial gaze bias results, a centerpiece of the paper, and presumably little effect on the cumulative gaze-time bias results.
Overall, these are important results, as they establish that the link between gaze and choice is not unique to humans. It also sets the stage for future neurophysiological studies to explore the neural mechanisms underlying behavior.
-
Reviewer #3 (Public Review):
Choosing between different options is a central aspect of behavior, and is not well understood. The pervasive influence of the neural reward machinery in determining behavior makes understanding such choices of central importance. The finding that monkeys exhibit the same choice biases linked to gaze behavior that humans do is a useful and necessary precursor to investigation of the neural mechanisms underlying choices of this kind. The authors find that monkeys are biased to choose the first option they look at, as well as the one they spend most time viewing, independent of the relative value of the choice. In addition, the animals tend to choose the last option they look at. The choice of targets that are not immediately discriminable in peripheral vision allows a clear interpretation of the results. The …
Reviewer #3 (Public Review):
Choosing between different options is a central aspect of behavior, and is not well understood. The pervasive influence of the neural reward machinery in determining behavior makes understanding such choices of central importance. The finding that monkeys exhibit the same choice biases linked to gaze behavior that humans do is a useful and necessary precursor to investigation of the neural mechanisms underlying choices of this kind. The authors find that monkeys are biased to choose the first option they look at, as well as the one they spend most time viewing, independent of the relative value of the choice. In addition, the animals tend to choose the last option they look at. The choice of targets that are not immediately discriminable in peripheral vision allows a clear interpretation of the results. The experiments were carefully conceived and executed, the analyses rigorous, and the logic of the conclusions is soundly based. Some more detail about the fixation behavior and fixation durations, not linked to the particular hypotheses tested, might have been useful.
-
