Sporulation activated via σ W protects Bacillus from a Tse1 peptidoglycan hydrolase T6SS effector
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
This manuscript will be of interest to readers in the field of microbiology, particularly those interested in bacterial interactions, secretion systems, and stress responses. It identifies the molecules and mechanisms that explain a competitive interaction between two soil-dwelling bacterial species. The data support most of the conclusions of the manuscript, but some controls are lacking and some of the interpretations are not fully justified by the experiments shown.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Bacterial communities constantly interact with various community members employing diverse offensive and defensive tools to reach coexistence. The extracellular matrix and sporulation are defensive mechanisms used by Bacillus cells when they interact with Pseudomonas strains expressing a type VI secretion system (T6SS). Here, we define Tse1 as the main toxin mobilized by the Pseudomonas T6SS that triggers sporulation in Bacillus . We characterize Tse1 as a peptidoglycan hydrolase that indirectly alters the dynamics and functionality of the Bacillus cell membrane. We also delineate the response of Bacillus cells to Tse1, which through the coordinated actions of the extracellular sigma factor σ W and the cytoplasmic histidine kinases KinA and KinB, culminates in activation of the sporulation cascade. We propose that this cellular developmental response is conserved in Bacilli to defend against the toxicity of T6SS-mobilized Tse1 effector.
Article activity feed
-

Author Response
Reviewer #1 (Public Review):
This work focuses on the mechanisms that underlie a previous observation by the authors that the type VI secretion system (T6SS) of a Pseudomonas chlororaphis (Pchl) strain can induce sporulation in Bacillus subtilis (Bsub). The authors bioinformatically characterize the T6SS system in Pchl and identify all the core components of the T6SS, as well as 8 putative effectors and their domain structures. They then show that the Pchl T6SS, and in particular its effector Tse1, is necessary to induce sporulation in Bsub. They demonstrate that Tse1 has peptidoglycan hydrolase activity and causes cell wall and cell membrane defects in Bsub. Finally, the authors also study the signaling pathway in Bsub that leads to the induction of sporulation, and their data suggest that cell wall damage may lead …
Author Response
Reviewer #1 (Public Review):
This work focuses on the mechanisms that underlie a previous observation by the authors that the type VI secretion system (T6SS) of a Pseudomonas chlororaphis (Pchl) strain can induce sporulation in Bacillus subtilis (Bsub). The authors bioinformatically characterize the T6SS system in Pchl and identify all the core components of the T6SS, as well as 8 putative effectors and their domain structures. They then show that the Pchl T6SS, and in particular its effector Tse1, is necessary to induce sporulation in Bsub. They demonstrate that Tse1 has peptidoglycan hydrolase activity and causes cell wall and cell membrane defects in Bsub. Finally, the authors also study the signaling pathway in Bsub that leads to the induction of sporulation, and their data suggest that cell wall damage may lead to the degradation of the anti-sigma factor RsiW, leading to activation of the extracellular sigma factor σW that causes increased levels of ppGpp. Sensing of high ppGpp levels by the kinases KinA and KinB may lead to phosphorylation of Spo0F, and induction of the sporulation cascade.
The findings add to the field's understanding of how competitive bacterial interactions work mechanistically and provide a detailed example of how bacteria may antagonize their neighbors, how this antagonism may be sensed, and the resulting defensive measures initiated.
While several of the conclusions of this paper are supported by the data, additional controls would bolster some aspects of the data, and some of the final interpretations are not substantiated by the current data.
- The Bsub signaling pathway that is proposed is intricate and extensive as shown in Fig 5A. However, the data supporting that is very sparse:
a) The authors show no data showing that the proteases PrsW and/or RasP, or the extracellular sigma factor σW are necessary, or that the cleavage of RsiW is needed, for induction of sporulation - this could presumably be tested using mutants of those genes.
It has been previously demonstrated that the proteases PrsW and/or RasP cleave RsiW under certain conditions such as alkaline-shock (Heinrich et al., 2009). In first place, PrsW cleaves RsiW and the resulting cleaved-RsiW serves as substrate to RasP. In the previous version of the manuscript, we already demonstrated that treatment with Tse1 causes damage to PG and delocalization of RsiW, however as the reviewer comments we did not show the participation of any of these proteases in the proposed signaling pathway. We have now generated single mutants in rsiW and prsW and they have been treated with Tse1. We have observed no variation in the levels of sporulation compared to untreated strains (Figure 1) a finding according to their suggested implication in the sporulation signaling pathway activated by Tse1. Positive controls, that is the single mutants grown at 37ºC, were still able to sporulate. This data has been added to Figure 6B in the new version of the manuscript.
As suggested by other reviewers, we have generated a sister plot of this figure showing the raw CFUs in each case. These data are included in Supplementary file 3. This experiment and the related figure have been incorporated into the new version of the manuscript.
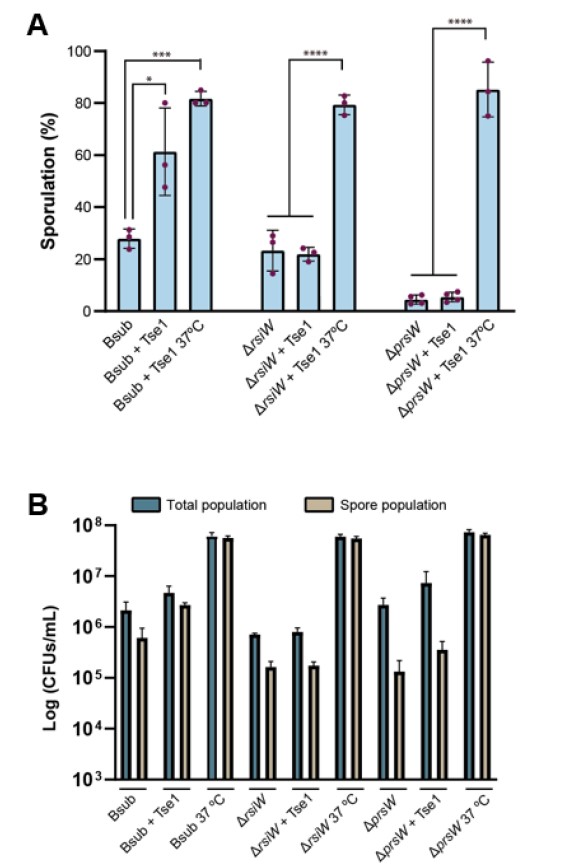
Figure 1. A) Quantification of the percentage of sporulated Bsub, rsiW and prsW cells after treatment with purified Tse1 showing that rsiW and prsW single mutants are blind to the presence of Tse1. B) Cell density (CFUs/mL) of total (blue bars) and sporulated population (brown bars) of different Bacillus strains (Bsub, ∆rsiW and ∆prsW) untreated and treated with Tse1. Sporulation at 37ºC is shown as positive control in each strain. Statistical significance was assessed via t-tests. *p value < 0.1, ***p value < 0.001, ****p value < 0.0001.
Similarly, they don't demonstrate that the levels of ppGpp increase in the cell upon exposure to Pchl.
We have not been able to measure the levels of ppGpp, however, given that in the same proposed sporulation cascade the levels of different nucleotides are altered (Kriel et al., 2013, Tojo et al., 2013, López and Kolter, 2010), we have alternatively analyzed the levels of ATP using an ATP Determination Kit (Thermo, A22066). We have found that ATP levels increased by 3-fold in Bsub cells treated with Tse1 compared to untreated control cells. Consistently, no increase in ATP levels were observed in rsiW or prsW mutants treated with Tse1. We have incorporated all the raw luminescence data obtained for each sample and treatment in Figure 6-source data 1. This experiment, figures (Figure 6A in the new version of the manuscript) and description in “Materials and Methods” have been added to the new version of the manuscript.
c) There is some data showing that kinA and kinB mutants don't induce sporulation (Fig supplement 7A), but that is lacking the 'no attacker' control that would demonstrate an induction.
We have included in the new version of the manuscript the ‘no attacker’ control sporulation (%). The figure shows that the presence of Pchl strains induces the sporulation of all kinase mutants. This new data has been incorporated in Figure 6-figure supplement 1A in the new version of the manuscript.
d) There is some data showing that RsiW may be cleaved (Fig 5C, D), but that data would benefit from a positive control showing that the lack of YFP foci is seen in a condition where RsiW is known to be cleaved, as well as from a time-course showing that the foci are present prior to the addition of Tse1, and then disappear. As it is shown now, it is possible that the addition of Tse1 just blocks the production of RsiW or its insertion into the membrane (especially given the membrane damage seen). Further, there is no data that the disappearance of the YFP loci requires the proteases PrsW and /or RasP - such data would also support the idea that the disappearance is due to cleavage of RsiW.
Thank you for your useful suggestion. It is important to consider that we have not seen repression of the expression of genes that encode any of the two proteases on cells treated with Tse1 in our transcriptomics analysis. However, we agree that additional experiments would enhance the significance of our findings. We have repeated the whole experiment including a positive control to demonstrate that YFP foci disappears in a condition in which RsiW is known to be degraded by PrsW and RasP. Bacillus cells have been incubated in medium at pH 10 which provokes an alkaline shock that triggers RsiW cleavage (Asai, 2017; Heinrich et al., 2009). As shown in Fiugre 6D under this condition we also observed disappearance of YFP foci . We have also provided extra images with quantification of average signal from YFP-foci in Figure 6-figure supplement 2 .
- The entire manuscript suggests that T6SS is solely responsible for the induction of sporulation. While T6SS does appear to play a major part in explaining the sporulation induction seen, in the absence of 'no attacker' controls for Fig. 2A, it is impossible to see this. From the data shown in Fig. 2C, and figure supplement 2A, the 'no attacker' sporulation rate seems to be ~20%, while the rate is ~40% with Pchl strains lacking T6SS, suggesting that an additional factor may be playing a role.
This must be a misunderstanding of the message of this manuscript. The conceptual fundament of this study was settled in our previous manuscript (Molina-Santiago et al., 2019). We demonstrated that B. subtilis sporulated in the presence of P. chlororaphis. Interestingly, the overgrowth of P. chlororaphis over B. subtilis colony did not eliminate cells of B. subtilis, given that most of them were sporulated. The data we obtained strongly suggested that a functional T6SS was involved in the cellular response of Bacillus in the close cell to cell contact. In this new manuscript, we have explored this idea, and found that indeed, the T6SS of P. chlororaphis mobilized at least one effector, Tse1, which is able to trigger sporulation in Bacillus. Thus we did not conclude, and neither have done in this new study, that T6SS is the only factor expressed by P. chlororaphis responsible for sporulation activation in Bacillus. We have accordingly rephrased some sentences of the manuscript to clarify the proposed implication of T6SS in B. subtilis sporulation.
In addition, as mentioned above, we have included data of sporulation percentages in the absence of an attacker to better compare the induction of sporulation observed in the presence of the different Pchl strains and in the presence of Tse1.
Reviewer #2 (Public Review):
In a previous study, the authors showed that cell-cell contact with Pseudomonas chlororaphis induces sporulation in Bacillus subtilis. Here, the authors build on this finding and elucidate the mechanism behind this observation. They describe the enzymatic activity of a protein (Tse1) secreted by the type VI secretion system (T6SS) of P. chlororaphis (Pch), which partially degrades the peptidoglycan (PG) of targeted B. subtilis cells and triggers a signal cascade culminating in sporulation.
Most of the key conclusions of this paper (Tse1 being secreted by the T6SS and inducing sporulation in targeted cells) are well supported by the data. One conclusion (sporulation response being an anti-T6SS "defense" strategy) is not well supported by the data and should be removed or rephrased.
The authors elucidate the enzymatic activity of Tse1, a T6SS effector protein, in a genus (Pseudomonas) of great interest to microbiologists, and to researchers studying the T6SS specifically. They also carefully dissect the cellular response (signal cascade and sporulation) of an important model organism (B. subtilis; Bsub) specifically to exposure to Tse1. The results describing this cellular response contribute substantially to our understanding of how T6SS effector proteins interact with cells of Gram-positive species.
My only major concerns regard the interpretation of these results as sporulation being an adaptive and/or specific response to attacks by the T6SS. I outline my reasoning below.
- Interpretation of sporulation as a "defense" mechanism/strategy against the T6SS. In order for a phenotype X to be regarded as a "defense against Y" mechanism, it has to be shown that phenotype X (sporulation in response to Tse1) evolved - at least in part - for the purposes of increasing survival in the presence of Y (T6SS attacker). There are no experiments in this study comparing e.g. a sporulating Bsub with a non-sporulating Bsub, that would allow testing if sporulation increases survival. The experiments carefully describe the cellular response to Tse1, but no inference can be made with regards to this being adaptive for Bsub, or if it helps the cells survive against T6SS attacks, etc. A more parsimonious explanation would be that Tse1 happens to target the PG and causes envelope stress, triggering sporulation. So, it would be a general stress response that also happens to be triggered by T6SS. Now, some general (cell envelope) stress responses are known to be very effective at protecting against the T6SS. But in those instances, a beneficial effect for survival in the face of T6SS attacks has been shown in dedicated experiments. Purely observing a response to a T6SS effector, as this study does (very well), is not evidence that the response has evolved for the purpose of surviving T6SS attacks. Tucked away in the supplement (and briefly mentioned in the main text) is data on Bsub and Bacillus cereus, showing that i) cell densities of the sporulating Bsub and a sporulating B. cereus strain are not affected by an active T6SS, and ii) cell densities of an asporogenic B. cereus are slightly reduced by an active T6SS. However, the effect sizes of density reduction by the T6SS in the asporogenic B. cereus are minute (20x10^6 vs. ~50x10^6). In typical killing assays against e.g. gram-negative strains, a typical effect size for T6SS killing would be a several order of magnitude reduction in survival of the target strain when exposed to a T6SS attacker. Based on this dataset alone (Figure Suppl. 8), I would say that all three Bacillus strains are not experiencing any "fitness-relevant" killing by the T6SS, which is in line with the T6SS often being useless against gram-positives when it comes to killing. Hence, no claims about fitness benefits of sporulation in response to a T6SS attack, or this being a "defense mechanism/strategy" should be made in the manuscript.
Thanks for this interesting introductory and specific comments. We agree with the reviewer and have rephrased some sentences of the manuscript. Sporulation is not an adaptive or specific response of Bacillus to T6SS, indeed and as stated by reviewer 2, sporulation is a general stress response. It might happen that the way the manuscript was written, at some points, gave the wrong impression. In consequence we have rephrased some sentences. Nevertheless, in Figure supplement 8 (in the new version of the manuscript is Figure 6-figure supplement 3) we made a mistake during generation of the Figure. We have again done this experiment and we have generated a new and corrected chart that shows three orders of magnitude reduction in survival of the asporogenic B. cereus strain in competition with Pchl mutant strains compared to Pchl WT strain. These new findings show that the absence of sporulation ability leads to a severe reduction in survival of Bacillus cereus DSM 2302 population in competition with Pchl with an active T6SS compared to the survival in competition with Pchl hcp mutant. In this figure, it is also shown that Bacillus population also decreased in competition with tse1 mutant, demonstrating that Tse1 is responsible for killing Bacillus. However, there is a statistical difference in the survival of Bacillus competing with hcp or tse1 mutants. The increased survival of Bacillus in the interaction with tse1 strain compared to Bacillus-hcp competition, is suggestive of the ability of this strain to deliver additional T6SS-dependent toxins. This observation is in accordance to the data presented in Fig. 2B, which indicated that tse1 mutant has an active T6SS able to kill E. coli.
- Data supporting baseline "no competitor" sporulation rates being no different from those triggered by T6SS mutants is not convincing. For the data shown in Fig. 2A, a key comparison here would be to show baseline Bsub sporulation rates in absence of a competitor. This measurement is shown in Fig supplement 2A, and the value shown there (roughly 22% on average) appears to be much lower than the average T6SS mutant shown in Fig. 2A. The main text states that sporulation rates induced rate by the different T6SS mutants are "statistically" similar to the no-competitor baseline (L206/207). I am not convinced by this, since i) overall sporulation rates (incl of WT Pch) appear to have been lower in the experiment shown in supplement 2A, so a direct comparison between the no-competitor baseline and the data shown in Fig. 2A is not possible; and ii) hcp and tse1 mutants were tested in different experiments throughout the study, and sporulation rates appear to consistently hover around 30-40%, which is higher than the roughly 22% for "no competitor" depicted in Supplement Fig2A. I am focussing on this, because for the interpretation of the results, and the main narrative of the paper, knowing if "simply interacting with a T6SS-negative P. chlororaphis" induces some sporulation would make a big difference. One sentence in the discussion adds to my confusion about this: L464/465, "... a strain lacking paar (Δpaar) had an active T6SS that triggered sporulation comparably to Δhcp, ΔtssA, and Δtse1 strains", suggesting that the authors' claims that even strains lacking active T6SS trigger increased sporulation (which I would agree with, based on the data).
We understand the reviewer's comment that a direct comparison between the two figures is not correct due to fluctuations of the baseline sporulation rates between experiments. To solve this issue, we have added the baseline "no competitor" sporulation percentages in the experiments represented in Figure 2B in the new version of the manuscript.
Related with the sporulation provoked by a T6SS-negative P. chlororaphis, the reviewer is right. Bacillus sporulation occurs due to many external factors (abiotic and biotic stresses) so the presence of P. chlororaphis in the competition already has an effect on the sporulation percentage of B. subtilis. Accordingly, we have removed the statement on the sporulation rates induced by the different T6SS mutants are "statistically" similar to the no-competitor. However, our previous data (Molina-Santiago, Nat Comm 2019) and current findings convincedly demonstrate the relevance of the T6SS and, specifically the Tse1 toxin, in the induction of sporulation at least in the close cell to cell contact.
- Claim regarding "bacteriolytic activity" when tse1 is heterologously expressed in E. coli. The data supporting this claim (Fig2-supplement 2C) only shows a lower net population growth rate after induction of tse1 (truncated vs. non-truncated) expression. This could be caused by: slower growth (but no death), equal growth (with some death), or a combination of the two. The claim of "bacteriolytic" activity in E. coli is therefore not supported by this dataset.
We agree with the reviewer and we have decided to remove this figure and the experiment of “bacteriolytic activity” given that it does not contribute conceptually to the message of the manuscript.
I cannot comment in more detail on the validity of the biochemistry/enzymatic activity assays as these are not my area of expertise.
Reviewer #3 (Public Review):
The authors identify tse1, a gene located in the type 6 secretion system (T6SS) locus of the bacterium Pseudomonas chlororaphis, as necessary and sufficient for induction of Bacillus subtilis sporulation. The authors demonstrate that Tse1 is a hydrolase that targets peptidoglycan in the bacterial cell wall, triggering activation of the regulatory sigma factor sigma-w. The sporulation-inducing effects of sigma-w are dependent on the downstream presence of the sensor histidine kinases KinA and KinB. Overall, this is a well-structured paper that uses a combination of methods including bacterial genetics, HPCL, microscopy, and immunohistochemistry to elucidate the mechanism of action of Tse1 against B. subtilis peptidoglycan. There are some concerns regarding a few experimental controls that were not included/discussed and (in a few figures) the visual representation of the data could be improved. The structure of the manuscript and experiments is such that key questions are addressed in a logical flow that demonstrates the mechanisms described by the authors.
To begin, we have concerns regarding the sporulation assays and their results. The data should be presented as "Percent sporulation" or "Sporulation (%)" - not as a "sporulation rate": there is no kinetic element to any of these measurements, so no rate is being measured (be careful of this in the text as well, for instance near lines 204). More importantly, there is no data provided to indicate that changes in percent spores are not instead just the death of non-sporulated cells. For example, imagine that within a population of B. subtilis cells, 85% of the cells are vegetative and 15% are spores. If, upon exposure to tse1, a large proportion of the vegetative cells are killed (say, 80% of them), this could lead to an apparent increase in sporulation: from 15% for the untreated population to ~50% of the treated, but the difference would be entirely due to a change in the vegetative population, not due to a change in sporulation. The authors need to clearly describe how they conducted their sporulation assays (currently there is no information about this in the methods) as well as provide the raw data of the counts of vegetative cells for their assays to eliminate this concern.
Thanks for the suggestion. We have changed all the titles and data presented as “sporulation rate” by “sporulation (%)” or “sporulation percentage”. As also suggested by reviewer 2, we have included the raw data of the CFUs counts of total population and sporulated cells to show that there is no substantial change in the rate of death. Also, we have added a section in Material and Methods to specify how sporulation assays have been done. Quote text:
“Sporulation assays
Spots of bacteria were resuspended in 1 mL sterile distilled water. Then, serial dilutions were made and cultured in LB solid media for vegetative cells CFU counts. The same serial dilutions were further heated at 80ºC for 10 minutes to kill vegetative cells and immediately cultured again in LB solid media. Plates were grown overnight at 28 ºC and the resulting colonies were counted to calculate the percentage of Bsub sporulation (%). A list of raw CFUs (total and spore population) from all figures with sporulation percentage is shown in Supplementary file 3.”
A related concern is regarding the analysis of the kinases and the effects of their deletions on the impact of Tse1. Previous literature shows that the basal levels of sporulation in a B. subtilis kinA or a kinB mutant are severely defective relative to a wild-type strain; these mutants sporulate poorly on their own. Therefore, the data presented on Lines 394+ and the associated Supplemental Figure regarding the sporulation defects of these two mutants are not compelling for showing that these kinases are required for this effector to act. It is likely that simply missing these kinases would severely impact the ability of these strains to sporulate at all, irrespective of the presence of Tse1, and no discussion of this confounding concern is discussed.
Previous literature shows that mutation of kinases affects sporulation of B. subtilis. Histidine kinases KinA and KinB are the first responsible for initiation of sporulation cascade upon phosphorylation of spo0F. However, as shown in Figure 6-figure supplement 1A, single mutants in these kinases (ΔkinA, ΔkinB) still sporulate given that the phosphorylation cascade is controlled by numerous intermediaries and other histidine kinases that form a multicomponent phosphorelay (KinA-E). In this context, the sporulation of B. subtilis can be also triggered by KinC or KinD in the absence of KinA or KinB, as KinC/KinD can act directly on the master regulator of sporulation Spo0A (Burbulys et al., 1991; Wang et al., 2017).
In addition, as suggested by reviewer 1, we have added to Figure 6-figure supplement 1A of the new version of the manuscript, the sporulation percentage 'no competitor' control of each kinase mutant and B. subtilis WT. The results show that, as commented by the reviewer and also supported by literature, these mutants sporulate poorly on their own in the absence of an attacker (none). However, as shown in the figure, all kinase mutants increase the sporulation percentage in the presence of a competitor.
Another concern is regarding the statistical tests used in Figure 2. For statistical tests in A, B, and D, it should be stated whether a post-test was used to correct for multiple comparisons, and, if so, which post-test was used. to provide a stronger control comparison. For C, we suggest the inclusion of a mock control in addition to the two conditions already included (i.e., an extraction from an E. coli strain expressing the empty vector)
We have clarified the statistical tests used in Figure 2. Briefly, we have used one-way ANOVA followed by the Dunnett test in Figure 2A, B and D for the statistical analysis of the sporulation percentage of Bsub in competition with Pchl as control group. In relation to Figure 2C, it is not possible to add a mock control with a strain carrying the empty vector, because this is a suicide plasmid (pDEST17) unable to replicate in E. coli without chromosome integration.
An additional concern regarding controls is that there is an absence of loading controls for the immunoblot assays. In Figure 5D and all immunoblot assays, there is no mention of a loading control, which is a critical control that should be included.
In the previous version of the manuscript, we already included a loading control for Figure 5D in Figure supplement 7B, both for cell and for supernatant fractions. In the new version of the manuscript, the loading control of Figure 6E (in the previous version of the manuscript Figure 5D) is shown in Figure 6-figure supplement 2C. We have also included the original unedited gels and blot (Figure 6-figure supplement 2- source data 1 and Figure 6-figure supplement 2-source data 2).
Some of the visualizations could be improved to help the reader understand and appropriately interpret the data presented. For instance, in Figures 3 and 4 the scale bars are different across each of the Figure's imaging panels. These should be scaled consistently for better comparison. Additionally, the red false colorization makes the printed images difficult to see. Black-and-white would be easier to see and would not subtract from the images.
The reviewer is right. Scales bar equal 2 in Figure 3A, but the length of the bars was not the same. We have edited the images to have the same magnifications for better comparison.
In relation to Figure 4, we have changed the magnifications and now all the figures have the same scale bars and magnifications. In addition, we have added more images of broader fields in Figure 4-figure supplement 1 which were used to measure the percentage of permeabilized cells and to obtain the fluorescence intensity measures shown in Figure 4.
An additional weakness of the paper is that the RNA-seq data is not fully investigated, and there is an absence of methods included regarding the RNA-seq differential abundance analysis (it is mentioned on L379-380 but no information is provided in the methods). As stated by the authors, 58% of differentially regulated genes belonged to the sw regulon, but the other 42% of genes are not discussed, and will hopefully be a target of future investigations.
The methods section has been modified for a better explanation of the RNA-seq differential abundance analysis. Quote text: “The raw reads were pre-processed with SeqTrimNext (Falgueras et al., 2010) using the specific NGS technology configuration parameters. This pre-processing removes low-quality, ambiguous and low-complexity stretches, linkers, adapters, vector fragments, and contaminated sequences while keeping the longest informative parts of the reads. SeqTrimNext also discarded sequences below 25 bp. Subsequently, clean reads were aligned and annotated using the Bsub reference genome with Bowtie2 (Langmead and Salzberg, 2012) in BAM files, which were then sorted and indexed using SAMtools v1.484(Li et al., 2009). Uniquely localized reads were used to calculate the read number value for each gene via Sam2counts (https://github.com/vsbuffalo/sam2counts). Differentially expressed genes (DEGs) were analyzed via DEgenes Hunter, which provides a combined p value calculated (based on Fisher’s method) using the nominal p values provided by edgeR (Robinson et al., 2010) and DEseq2. This combined p value was adjusted using the Benjamini-Hochberg (BH) procedure (false discovery rate approach) and used to rank all the obtained DEGs. For each gene, combined p value < 0.05 and log2-fold change > 1 or < −1 were considered as the significance threshold”
Regarding the RNA-seq analysis, we are aware of the amount of information that can be extracted. Previous to filtering the information shown in the manuscript, we have done bioinformatic analysis trying to find a connection with the cellular response, that is increase of sporulation. Besides this, we had some observations but with no direct connection to sporulation, which would be interesting to pursue in future studies, but not for the clarity of this story (Figure 23 below). In any case, we are including the whole picture of the transcriptomics changes occurring in Bsub after treatment with Tse1. KEGG pathway analyses of genes differentially expressed showed induction of flagellar assembly and aminobenzoate degradation, nitrogen and amino acid metabolisms. Interestingly, fatty acid degradation and CAMP resistance pathways were also induced, probably related to changes suffered in the cell wall after the action of Tse1 toxin. On the other hand, synthesis and degradation of ketone bodies pathway was mostly repressed.
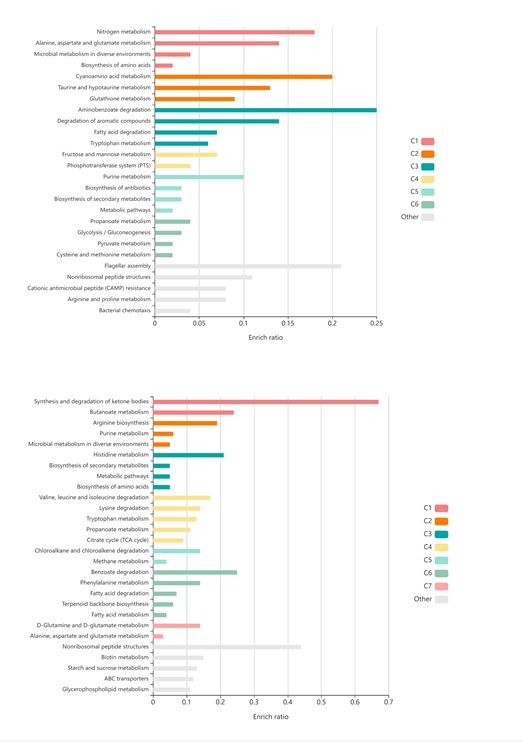
Figure 2. KEGG pathway analyses of genes differentially expressed occurring in Bsub after treatment with Tse1.
Another methodological concern in this paper is the limited details provided for the calculation of the permeabilization rate (Figure 4, L359, L662-664). It is not clear how, or if, cell density was controlled for in these experiments.
We agree with the reviewer and we have explained with more detail how the permeabilization rate was calculated. Quote text: “N=3 for Bsub treated with Tse1 and N=3 for untreated Bsub. N refers to the number of CLSM fields analyzed to calculate the number of permeabilized cells of the total of cells in the field”
Finally, one weakness of the paper is the broad conclusions that they draw. The authors claim that the mechanism of sporulation activation is conserved across Bacilli when the authors only test one B. subtilis and one B. cereus strain. They further argue (lines 469+) that Tse1 requires a PAAR repeat for its targeting, but do not provide direct evidence for this possibility.
We have reduced the tone of the final conclusion in order to specify that the activation of sporulation is a mechanism that can be found in different Bacillus species such as Bsub and Bcer. Related with the second appreciation, we have included a further explanation for this argument. Quote text: “As shown in Figure 2B, a paar mutant has an active T6SS able to kill E. coli. However, as shown in Figure 2A, we noticed that a paar mutant (which encodes tse1) is not able to trigger B. subtilis sporulation to a similar level than Pchl WT strain. Given that paar deletion apparently abolishes Tse1 secretion, we suggest that Tse1 is a PAAR-associated effector that requires a PAAR repeat domain protein to be targeted for secretion, thereby increasing Bacillus sporulation during contact with Pseudomonas cells (Cianfanelli et al., 2016; Hachani et al., 2014; Whitney et al., 2014)”.
-
Evaluation Summary:
This manuscript will be of interest to readers in the field of microbiology, particularly those interested in bacterial interactions, secretion systems, and stress responses. It identifies the molecules and mechanisms that explain a competitive interaction between two soil-dwelling bacterial species. The data support most of the conclusions of the manuscript, but some controls are lacking and some of the interpretations are not fully justified by the experiments shown.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
-
Reviewer #1 (Public Review):
This work focuses on the mechanisms that underlie a previous observation by the authors that the type VI secretion system (T6SS) of a Pseudomonas chlororaphis (Pchl) strain can induce sporulation in Bacillus subtilis (Bsub). The authors bioinformatically characterize the T6SS system in Pchl and identify all the core components of the T6SS, as well as 8 putative effectors and their domain structures. They then show that the Pchl T6SS, and in particular its effector Tse1, is necessary to induce sporulation in Bsub. They demonstrate that Tse1 has peptidoglycan hydrolase activity and causes cell wall and cell membrane defects in Bsub. Finally, the authors also study the signaling pathway in Bsub that leads to the induction of sporulation, and their data suggest that cell wall damage may lead to the degradation …
Reviewer #1 (Public Review):
This work focuses on the mechanisms that underlie a previous observation by the authors that the type VI secretion system (T6SS) of a Pseudomonas chlororaphis (Pchl) strain can induce sporulation in Bacillus subtilis (Bsub). The authors bioinformatically characterize the T6SS system in Pchl and identify all the core components of the T6SS, as well as 8 putative effectors and their domain structures. They then show that the Pchl T6SS, and in particular its effector Tse1, is necessary to induce sporulation in Bsub. They demonstrate that Tse1 has peptidoglycan hydrolase activity and causes cell wall and cell membrane defects in Bsub. Finally, the authors also study the signaling pathway in Bsub that leads to the induction of sporulation, and their data suggest that cell wall damage may lead to the degradation of the anti-sigma factor RsiW, leading to activation of the extracellular sigma factor σW that causes increased levels of ppGpp. Sensing of high ppGpp levels by the kinases KinA and KinB may lead to phosphorylation of Spo0F, and induction of the sporulation cascade.
The findings add to the field's understanding of how competitive bacterial interactions work mechanistically and provide a detailed example of how bacteria may antagonize their neighbors, how this antagonism may be sensed, and the resulting defensive measures initiated.
While several of the conclusions of this paper are supported by the data, additional controls would bolster some aspects of the data, and some of the final interpretations are not substantiated by the current data.
- The Bsub signaling pathway that is proposed is intricate and extensive as shown in Fig 5A. However, the data supporting that is very sparse:
a) The authors show no data showing that the proteases PrsW and/or RasP, or the extracellular sigma factor σW are necessary, or that the cleavage of RsiW is needed, for induction of sporulation - this could presumably be tested using mutants of those genes.
b) Similarly, they don't demonstrate that the levels of ppGpp increase in the cell upon exposure to Pchl.
c) There is some data showing that kinA and kinB mutants don't induce sporulation (Fig supplement 7A), but that is lacking the 'no attacker' control that would demonstrate an induction.
d) There is some data showing that RsiW may be cleaved (Fig 5C, D), but that data would benefit from a positive control showing that the lack of YFP foci is seen in a condition where RsiW is known to be cleaved, as well as from a time-course showing that the foci are present prior to the addition of Tse1, and then disappear. As it is shown now, it is possible that the addition of Tse1 just blocks the production of RsiW or its insertion into the membrane (especially given the membrane damage seen). Further, there is no data that the disappearance of the YFP loci requires the proteases PrsW and /or RasP - such data would also support the idea that the disappearance is due to cleavage of RsiW.
- The entire manuscript suggests that T6SS is solely responsible for the induction of sporulation. While T6SS does appear to play a major part in explaining the sporulation induction seen, in the absence of 'no attacker' controls for Fig. 2A, it is impossible to see this. From the data shown in Fig. 2C, and figure supplement 2A, the 'no attacker' sporulation rate seems to be ~20%, while the rate is ~40% with Pchl strains lacking T6SS, suggesting that an additional factor may be playing a role. -
Reviewer #2 (Public Review):
In a previous study, the authors showed that cell-cell contact with Pseudomonas chlororaphis induces sporulation in Bacillus subtilis. Here, the authors build on this finding and elucidate the mechanism behind this observation. They describe the enzymatic activity of a protein (Tse1) secreted by the type VI secretion system (T6SS) of P. chlororaphis (Pch), which partially degrades the peptidoglycan (PG) of targeted B. subtilis cells and triggers a signal cascade culminating in sporulation.
Most of the key conclusions of this paper (Tse1 being secreted by the T6SS and inducing sporulation in targeted cells) are well supported by the data. One conclusion (sporulation response being an anti-T6SS "defense" strategy) is not well supported by the data and should be removed or rephrased.
The authors elucidate the …
Reviewer #2 (Public Review):
In a previous study, the authors showed that cell-cell contact with Pseudomonas chlororaphis induces sporulation in Bacillus subtilis. Here, the authors build on this finding and elucidate the mechanism behind this observation. They describe the enzymatic activity of a protein (Tse1) secreted by the type VI secretion system (T6SS) of P. chlororaphis (Pch), which partially degrades the peptidoglycan (PG) of targeted B. subtilis cells and triggers a signal cascade culminating in sporulation.
Most of the key conclusions of this paper (Tse1 being secreted by the T6SS and inducing sporulation in targeted cells) are well supported by the data. One conclusion (sporulation response being an anti-T6SS "defense" strategy) is not well supported by the data and should be removed or rephrased.
The authors elucidate the enzymatic activity of Tse1, a T6SS effector protein, in a genus (Pseudomonas) of great interest to microbiologists, and to researchers studying the T6SS specifically. They also carefully dissect the cellular response (signal cascade and sporulation) of an important model organism (B. subtilis; Bsub) specifically to exposure to Tse1. The results describing this cellular response contribute substantially to our understanding of how T6SS effector proteins interact with cells of Gram-positive species.
My only major concerns regard the interpretation of these results as sporulation being an adaptive and/or specific response to attacks by the T6SS. I outline my reasoning below.
- Interpretation of sporulation as a "defense" mechanism/strategy against the T6SS. In order for a phenotype X to be regarded as a "defense against Y" mechanism, it has to be shown that phenotype X (sporulation in response to Tse1) evolved - at least in part - for the purposes of increasing survival in the presence of Y (T6SS attacker). There are no experiments in this study comparing e.g. a sporulating Bsub with a non-sporulating Bsub, that would allow testing if sporulation increases survival. The experiments carefully describe the cellular response to Tse1, but no inference can be made with regards to this being adaptive for Bsub, or if it helps the cells survive against T6SS attacks, etc. A more parsimonious explanation would be that Tse1 happens to target the PG and causes envelope stress, triggering sporulation. So, it would be a general stress response that also happens to be triggered by T6SS. Now, some general (cell envelope) stress responses are known to be very effective at protecting against the T6SS. But in those instances, a beneficial effect for survival in the face of T6SS attacks has been shown in dedicated experiments. Purely observing a response to a T6SS effector, as this study does (very well), is not evidence that the response has evolved for the purpose of surviving T6SS attacks. Tucked away in the supplement (and briefly mentioned in the main text) is data on Bsub and Bacillus cereus, showing that i) cell densities of the sporulating Bsub and a sporulating B. cereus strain are not affected by an active T6SS, and ii) cell densities of an asporogenic B. cereus are slightly reduced by an active T6SS. However, the effect sizes of density reduction by the T6SS in the asporogenic B. cereus are minute (20x10^6 vs. ~50x10^6). In typical killing assays against e.g. gram-negative strains, a typical effect size for T6SS killing would be a several order of magnitude reduction in survival of the target strain when exposed to a T6SS attacker. Based on this dataset alone (Figure Suppl. 8), I would say that all three Bacillus strains are not experiencing any "fitness-relevant" killing by the T6SS, which is in line with the T6SS often being useless against gram-positives when it comes to killing. Hence, no claims about fitness benefits of sporulation in response to a T6SS attack, or this being a "defense mechanism/strategy" should be made in the manuscript.
- Data supporting baseline "no competitor" sporulation rates being no different from those triggered by T6SS mutants is not convincing. For the data shown in Fig. 2A, a key comparison here would be to show baseline Bsub sporulation rates in absence of a competitor. This measurement is shown in Fig supplement 2A, and the value shown there (roughly 22% on average) appears to be much lower than the average T6SS mutant shown in Fig. 2A. The main text states that sporulation rates induced by the different T6SS mutants are "statistically" similar to the no-competitor baseline (L206/207). I am not convinced by this, since i) overall sporulation rates (incl of WT Pch) appear to have been lower in the experiment shown in supplement 2A, so a direct comparison between the no-competitor baseline and the data shown in Fig. 2A is not possible; and ii) hcp and tse1 mutants were tested in different experiments throughout the study, and sporulation rates appear to consistently hover around 30-40%, which is higher than the roughly 22% for "no competitor" depicted in Supplement Fig2A. I am focussing on this, because for the interpretation of the results, and the main narrative of the paper, knowing if "simply interacting with a T6SS-negative P. chlororaphis" induces some sporulation would make a big difference. One sentence in the discussion adds to my confusion about this: L464/465, "... a strain lacking paar (Δpaar) had an active T6SS that triggered sporulation comparably to Δhcp, ΔtssA, and Δtse1 strains", suggesting that the authors' claims that even strains lacking active T6SS trigger increased sporulation (which I would agree with, based on the data).
- Claim regarding "bacteriolytic activity" when tse1 is heterologously expressed in E. coli. The data supporting this claim (Fig2-supplement 2C) only shows a lower net population growth rate after induction of tse1 (truncated vs. non-truncated) expression. This could be caused by: slower growth (but no death), equal growth (with some death), or a combination of the two. The claim of "bacteriolytic" activity in E. coli is therefore not supported by this dataset.
I cannot comment in more detail on the validity of the biochemistry/enzymatic activity assays as these are not my area of expertise.
-
Reviewer #3 (Public Review):
The authors identify tse1, a gene located in the type 6 secretion system (T6SS) locus of the bacterium Pseudomonas chlororaphis, as necessary and sufficient for induction of Bacillus subtilis sporulation. The authors demonstrate that Tse1 is a hydrolase that targets peptidoglycan in the bacterial cell wall, triggering activation of the regulatory sigma factor sigma-w. The sporulation-inducing effects of sigma-w are dependent on the downstream presence of the sensor histidine kinases KinA and KinB. Overall, this is a well-structured paper that uses a combination of methods including bacterial genetics, HPCL, microscopy, and immunohistochemistry to elucidate the mechanism of action of Tse1 against B. subtilis peptidoglycan. There are some concerns regarding a few experimental controls that were not …
Reviewer #3 (Public Review):
The authors identify tse1, a gene located in the type 6 secretion system (T6SS) locus of the bacterium Pseudomonas chlororaphis, as necessary and sufficient for induction of Bacillus subtilis sporulation. The authors demonstrate that Tse1 is a hydrolase that targets peptidoglycan in the bacterial cell wall, triggering activation of the regulatory sigma factor sigma-w. The sporulation-inducing effects of sigma-w are dependent on the downstream presence of the sensor histidine kinases KinA and KinB. Overall, this is a well-structured paper that uses a combination of methods including bacterial genetics, HPCL, microscopy, and immunohistochemistry to elucidate the mechanism of action of Tse1 against B. subtilis peptidoglycan. There are some concerns regarding a few experimental controls that were not included/discussed and (in a few figures) the visual representation of the data could be improved. The structure of the manuscript and experiments is such that key questions are addressed in a logical flow that demonstrates the mechanisms described by the authors.
To begin, we have concerns regarding the sporulation assays and their results. The data should be presented as "Percent sporulation" or "Sporulation (%)" - not as a "sporulation rate": there is no kinetic element to any of these measurements, so no rate is being measured (be careful of this in the text as well, for instance near lines 204). More importantly, there is no data provided to indicate that changes in percent spores are not instead just the death of non-sporulated cells. For example, imagine that within a population of B. subtilis cells, 85% of the cells are vegetative and 15% are spores. If, upon exposure to tse1, a large proportion of the vegetative cells are killed (say, 80% of them), this could lead to an apparent increase in sporulation: from 15% for the untreated population to ~50% of the treated, but the difference would be entirely due to a change in the vegetative population, not due to a change in sporulation. The authors need to clearly describe how they conducted their sporulation assays (currently there is no information about this in the methods) as well as provide the raw data of the counts of vegetative cells for their assays to eliminate this concern.
A related concern is regarding the analysis of the kinases and the effects of their deletions on the impact of Tse1. Previous literature shows that the basal levels of sporulation in a B. subtilis kinA or a kinB mutant are severely defective relative to a wild-type strain; these mutants sporulate poorly on their own. Therefore, the data presented on Lines 394+ and the associated Supplemental Figure regarding the sporulation defects of these two mutants are not compelling for showing that these kinases are required for this effector to act. It is likely that simply missing these kinases would severely impact the ability of these strains to sporulate at all, irrespective of the presence of Tse1, and no discussion of this confounding concern is discussed.
Another concern is regarding the statistical tests used in Figure 2. For statistical tests in A, B, and D, it should be stated whether a post-test was used to correct for multiple comparisons, and, if so, which post-test was used. For C, we suggest the inclusion of a mock control in addition to the two conditions already included (i.e., an extraction from an E. coli strain expressing the empty vector) to provide a stronger control comparison.
An additional concern regarding controls is that there is an absence of loading controls for the immunoblot assays. In Figure 5D and all immunoblot assays, there is no mention of a loading control, which is a critical control that should be included.
Some of the visualizations could be improved to help the reader understand and appropriately interpret the data presented. For instance, in Figures 3 and 4 the scale bars are different across each of the Figure's imaging panels. These should be scaled consistently for better comparison. Additionally, the red false colorization makes the printed images difficult to see. Black-and-white would be easier to see and would not subtract from the images.
An additional weakness of the paper is that the RNA-seq data is not fully investigated, and there is an absence of methods included regarding the RNA-seq differential abundance analysis (it is mentioned on L379-380 but no information is provided in the methods). As stated by the authors, 58% of differentially regulated genes belonged to the w regulon, but the other 42% of genes are not discussed, and will hopefully be a target of future investigations.
Another methodological concern in this paper is the limited details provided for the calculation of the permeabilization rate (Figure 4, L359, L662-664). It is not clear how, or if, cell density was controlled for in these experiments.
Finally, one weakness of the paper is the broad conclusions that they draw. The authors claim that the mechanism of sporulation activation is conserved across Bacilli when the authors only test one B. subtilis and one B. cereus strain. They further argue (lines 469+) that Tse1 requires a PAAR repeat for its targeting, but do not provide direct evidence for this possibility.
-
