Foveal vision anticipates defining features of eye movement targets
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
This study measures the strength, timing, feature specificity and for the first time - spatial extent - of pre-saccadic processing of peripheral target information at the fovea, when fixation is not constrained. The authors conclude that saccade preparation causes feature-specific pre-saccadic visual enhancement restricted largely to the center of gaze. The manuscript is based on a rigorously curated dataset, it is well written and illustrated, and will be of potential interest to a broad readership of vision scientists, neuroscientists, and modelers. However, some key weaknesses in the interpretation of data were identified, which call into question the conclusion that the foveal enhancement observed can only be explained in terms of saccade preparation, thus weakening our ability to infer its mechanistic substrates.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
High-acuity foveal processing is vital for human vision. Nonetheless, little is known about how the preparation of large-scale rapid eye movements (saccades) affects visual sensitivity in the center of gaze. Based on findings from passive fixation tasks, we hypothesized that during saccade preparation, foveal processing anticipates soon-to-be fixated visual features. Using a dynamic large-field noise paradigm, we indeed demonstrate that defining features of an eye movement target are enhanced in the pre-saccadic center of gaze. Enhancement manifested as higher Hit Rates for foveal probes with target-congruent orientation and a sensitization to incidental, target-like orientation information in foveally presented noise. Enhancement was spatially confined to the center of gaze and its immediate vicinity, even after parafoveal task performance had been raised to a foveal level. Moreover, foveal enhancement during saccade preparation was more pronounced and developed faster than enhancement during passive fixation. Based on these findings, we suggest a crucial contribution of foveal processing to trans-saccadic visual continuity: Foveal processing of saccade targets commences before the movement is executed and thereby enables a seamless transition once the center of gaze reaches the target.
Article activity feed
-

-

Author reponse
Reviewer #1 (Public Review):
In their paper, Kroell and Rolfs use a set of sophisticated psychophysical experiments in visually-intact observers, to show that visual processing at the fovea within the 250ms or so before saccading to a peripheral target containing orientation information, is influenced by orientation signals at the target. Their approach straddles the boundary between enforcing fixation throughout stimulus presentation (a standard in the field) and leaving it totally unconstrained. As such, they move the field of saccade pre-processing towards active vision in order to answer key questions about whether the fovea predicts features at the gaze target, over what time frame, with what precision, and over what spatial extent around the foveal center. The results support the notion that there is …
Author reponse
Reviewer #1 (Public Review):
In their paper, Kroell and Rolfs use a set of sophisticated psychophysical experiments in visually-intact observers, to show that visual processing at the fovea within the 250ms or so before saccading to a peripheral target containing orientation information, is influenced by orientation signals at the target. Their approach straddles the boundary between enforcing fixation throughout stimulus presentation (a standard in the field) and leaving it totally unconstrained. As such, they move the field of saccade pre-processing towards active vision in order to answer key questions about whether the fovea predicts features at the gaze target, over what time frame, with what precision, and over what spatial extent around the foveal center. The results support the notion that there is feature-selective enhancement centered on the center of gaze, rather than on the predictively remapped location of the target. The results further show that this enhancement extends about 3 deg radially from the foveal center and that it starts ~ 200ms or so before saccade onset. They also show that this enhancement is reinforced if the target remains present throughout the saccade. The hypothesized implications of these findings are that they could enable continuity of perception trans-saccadically and potentially, improve post-saccadic gaze correction.
Strengths:
The findings appear solid and backed up by converging evidence from several experimental manipulations. These included several approaches to overcome current methodological constraints to the critical examination of foveal processing while being careful not to interfere with saccade planning and performance. The authors examined the spatial frequency characteristics of the foveal enhancement relative, hit rates and false alarm rates for detecting a foveal probe that was congruent or incongruent in terms of orientation to the peripheral saccade target embedded in flickering, dynamic noise (i/f )images. While hit rates are relatively easy to interpret, the authors also reconstructed key features of the background noise to interpret false alarms as reflecting foveal enhancement that could be correlated with target orientation signals. The study also - in an extensive Supplementary Materials section - uses appropriate statistical analyses and controls for multiple factors impacting experimental/stimulus design and analysis. The approach, as well as the level of care towards experimental details provided in this manuscript, should prove welcome and useful for any other investigators interested in the questions posed.
Weaknesses:
I find no major weaknesses in the experiments, analyses or interpretations. The conclusions of the paper appear well supported by the data. My main suggestion would be to see a clearer discussion of the implications of the present findings for truly naturalistic, visually-guided performance and action. Please consider the implication of the phenomena and behaviors reported here when what is located at the gaze center (while peripheral targets are present), is not a noisy, relatively feature-poor, low-saliency background, but another high-saliency target, likely crowded by other nearby targets. As such, a key question that emerges and should be addressed in the Discussion at least is whether the fovea's role described in the present experiments is restricted to visual scenarios used here, or whether they generalize to the rather different visual environments of everyday life.
This is a very interesting question. While we cannot provide a definite answer, we have added a paragraph discussing the role of foveal prediction in more naturalistic visual contexts to the Discussion section (‘Does foveal prediction transfer to other visual features and complex natural environments?’). We pasted this paragraph in response to another comment in the ‘Recommendations for the authors’ section below. We suggest that “the pre-saccadic decrease in foveal sensitivity demonstrated previously[9] as well as in our own data (Figure 2B) may boost the relative strength of fed-back signals by reducing the conspicuity of foveal feedforward input”, presumably allowing the foveal prediction mechanism to generalize to more naturalistic environments with salient foveal stimulation.
Reviewer #2 (Public Review):
Human and primates move their eyes with rapid saccades to reposition the high-resolution region of the retina, the fovea, over objects of interest. Thus, each saccade involves moving the fovea from a pre-saccadic location to a saccade target. Although it has been long known that saccades profoundly alter visual processing at the time of saccade, scientists simply do not know how the brain combines information across saccades to support our normal perceptual experience. This paper addresses a piece of that puzzle by examining how eye movements affect processing at the fovea before it moves. Using a dynamic noise background and a dual psychophysical task, the authors probe both the performance and selectivity of visual processing for orientation at the fovea in the few hundred milliseconds preceding a saccade. They find that hit rates and false alarm rates are dynamically and automatically modulated by the saccade planning. By taking advantage of the specific sequence of noise shown on each trial, they demonstrate that the tuning of foveal processing is affected by the orientation of the saccade target suggesting foveal specific feedback.
A major strength of the paper is the experimental design. The use of dynamic filtered noise to probe perceptual processing is a clever way of measuring the dynamics of selectivity at the fovea during saccade preparation. The use of a dual-task allows the authors to evaluate the tuning of foveal processing as well and how it depends on the peripheral target orientation. They show compellingly that the orientation of the saccade target (the future location of the fovea) affects processing at the fovea before it moves.
There are two weaknesses with the paper in its current form. The first is that the key claim of foveal "enhancement" relies on the tuning of the false alarms. A more standard measure of enhancement would be to look at the sensitivity, or d-prime, of the performance on the task. In this study, hits and false alarms increase together, which is traditionally interpreted as a criterion shift and not an enhancement. However, because of the external noise, false alarms are driven by real signals. The authors are aware of this and argue that the fact that the false alarms are tuned indicates enhancement. But it is unclear to me that a criterion shift wouldn't also explain this tuning and the change in the noise images. For example, in a task with 4 alternative choices (Present/Congruent, Present/Incongruent, Absent/Congruent, Absent/Incongruent), shifting the criterion towards the congruent target would increase hits and false alarms for that target and still result in a tuned template (because that template is presumably what drove the decision variable that the adjusted criterion operates on). I believe this weakness could be addressed with a computational model that shows that a criterion shift on the output of a tuned template cannot produce the pattern of hits and false alarms.
We thank the reviewer for this comment. We will present three arguments, each of which suggests that our effects are perceptual in nature and cannot be explained by a shift in decision criterion: (1) the temporal specificity of the difference in Hit Rates (HRs), (2) the spatial specificity of the difference in HRs and (3) the phenomenological quality of the foveally predicted signal. In general, a criterion shift would indeed affect hits and false alarms alike. Nonetheless, the difference in HRs only manifested under specific and meaningful conditions:
First, the increase in congruent as compared to incongruent HRs, i.e., enhancement, was temporally specific: congruent and incongruent HRs were virtually identical when the probe appeared in a baseline time bin or one (Figure 2B) or even two (Figure 4A) early pre-saccadic time bins. Based on another reviewer’s comment, we collected additional data to measure the time course and extent of foveal enhancement during fixation. While pre-saccadic enhancement developed rapidly, enhancement started to emerge 200 ms after target onset during fixation. Crucially, these time courses mirror the typical temporal development of visual sensitivity during pre-saccadic attention shifts and covert attentional allocation, respectively[8,33]. We are unaware of data demonstrating similar temporal specificity for a shift in decision criterion. One could argue that a template of the target orientation needs to build up before it can influence criterion. Nonetheless, this template would be expected to remain effective after this initial temporal threshold has been crossed. In contrast, we observe pronounced enhancement in medium but not late stages of saccade preparation in the PRE-only condition (Figure 4A).
Second, it has been argued that a defining difference between innately perceptual effects and post-perceptual criterion shifts is their spatial specificity[53]: in opposition to perceptual effects, criterion shifts should manifest in a spatially global fashion. Due to a parafoveal control condition detailed in our reply to the next comment, we maintain the claim that enhancement is spatially specific: congruent HRs exceeded incongruent ones within a confined spatial region around the center of gaze. We did not observe enhancement for probes presented at 3 dva eccentricity even when we raised parafoveal performance to a foveal level by adaptively increasing probe contrast. The accuracy of saccade landing or, more specifically, the mean remapped target location (Figure 3B) influenced the spatial extent of the enhanced region in a fashion that is reconcilable with previous findings[30]. A criterion shift that is both spatially and temporally selective, follows the time course of pre-saccadic or covert attention depending on observers’ oculomotor behavior, does not remain effective throughout the entire trial after its onset, is sensitive to the mean remapped target location across trials, and does not apply to parafoveal probes even after their contrast has been increased to match foveal performance, would be unprecedented in the literature and, even if existent, appear just as functionally meaningful as sensitivity changes occurring under the same conditions.
Lastly and on a more informal note, we would like to describe a phenomenological percept that was spontaneously reported by 6 out of 7 observers in Experiment 1 and experienced by the author L.M.K. many times. On a small subset of trials, participants in our paradigms have the strong phenomenological impression of perceiving the target in the pre-saccadic center of gaze. This percept is rare but so pronounced that some observers interrupt the experiment to ask which probe orientation they should report if they had perceived two on the same trial (“The orientation of the normal probe or of the one that looked exactly like the target”). Interestingly, the actual saccade target and its foveal equivalent are perceived simultaneously in two spatiotopically separate locations, suggesting that this percept cannot be ascribed to a temporal misjudgment of saccade execution (after which the target would have actually been foveated). We have no data to prove this observation but nonetheless wanted to share it. Experiencing it ourselves has left us with no doubt that the fed-back signal is truly – and almost eerily – perceptual in nature.
The analysis suggested by the reviewer is very interesting. Yet for several reasons stated in the ‘Suggestions to the authors’ section, our dataset is not cut out for an analysis of noise properties at this level of complexity. We had always planned to resolve these concerns experimentally, i.e., by demonstrating specificity in HRs. We believe that our arguments above provide a strong case for a perceptual phenomenon and have incorporated them into the Discussion of our revised manuscript.
The second weakness is that the author's claim that feedback is spatially selective to the fovea is confounded by the fact that acuity and contrast sensitivity are higher in the fovea. Therefore, the subject's performance would already be spatially tuned. Even the very central degree, the foveola, is inhomogeneous. Thus, finding spatially-tuned sensitivity to the probes may simply indicate global feature gain on top of already spatially tuned processing in the fovea. Another possible explanation that is consistent with the "no enhancement" interpretation is that the fovea has increased. This is consistent with the observation that the congruency effects were aligned to the center of gaze and not the saccade endpoint. It looks from the Gaussian fits that a single gain parameter would explain the difference in the shape of the congruent and incongruent hit rates, but I could not figure out if this was explicitly tested from the existing methods. Additional experiments without prepared saccades would be an easy way to address this issue. Is the hit rate tuned when there is no saccade preparation? If so, it seems likely that the spatial selectivity is not tuned feedback, but inhomogeneous feedforward processing.
We fully agree. We do not consider a fixation condition diagnostic to resolve this question since, as of now, correlates of foveal feedback have exclusively been observed during fixation. In those studies, it was suggested that the effect, i.e., a foveal representation of peripheral stimuli, reflects the automatic preparation of an eye movement that was simply not executed[11,12,14]. To address another reviewer’s comment, we collected additional data in a fixation experiment. The probe stimulus could exclusively appear in the screen center (as in Experiment 1) and observers maintained fixation throughout the trial. While pre-saccadic congruency effects were significantly more pronounced and developed faster, congruency effects did emerge during fixation when the probe appeared 200 ms after the target. If pre-saccadic processes indeed spill over to fixation tasks to some extent and trigger relevant neural mechanisms even when no saccade is executed, we could expect a similar feedback-induced spatial profile during fixation. Since this matches the reviewer’s prediction if the pre-saccadic profiles resulted from inhomogeneous feedforward processing, we do not consider a fixation condition suitable to distinguish between both hypotheses.
To test whether the tuning of enhancement is effectively a consequence of declining visual performance in the parafovea/periphery, we instead raised parafoveal performance to a foveal level by adaptively increasing the opacity of the probe: while leaving all remaining experimental parameters unchanged, we presented the probe in one of two parafoveal locations, i.e., 3 dva to the left or right of the screen center. Observers were explicitly informed about the placement of the probe. We administered a staircase procedure to determine the probe opacity at which performance for parafoveal target-incongruent probes would be just as high as foveal performance had been in the preceding sessions. While the foveal probe was presented at a median opacity of 28.3±7.6%, a parafoveal opacity of 39.0±11.1% was required to achieve the same performance level. As a result, the gray dot at 0 dva in the figure below represents the incongruent HR in the center of gaze and ranges at 80% on the y-axis. The gray dots at ±3 dva represent incongruent parafoveal HRs and also range at ~80% on the y-axis. Using the reviewer’s terminology, we effectively removed the influence of acuity- (or contrast-sensitivity-) dependent spatial tuning. If the spatial profiles had indeed been the result of “global feature gain on top of already spatially tuned processing“, this manipulation should render parafoveal feature gain just as detectable as foveal feature gain. Instead, congruent and incongruent parafoveal HRs were statistically indistinguishable (away from the saccade target: p = .127, BF10 = 0.531; towards the saccade target: p = .336, BF10 = 0.352), inconsistent with the idea of a spatially global feature gain.
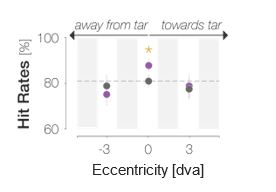
We had included these data in our initial submission. They were collected in the same observers that contributed the spatial profiles (Experiment 2). The data points at 0 dva in the reduced figure above correspond to the foveal probe location in Figure 2D. The data points at ±3 dva had been plotted and discussed in our initial submission, yet only very briefly. Based on this and another reviewer’s comment, we realize that we should have explained this condition more extensively in the main text rather than in the Methods and have added a dedicated paragraph to the Results section.
This paper is important because it compellingly demonstrates that visual processing in the fovea anticipates what is coming once the eyes move. The exact form of the modulation remains unclear and the authors could do more to support their interpretations. However, understanding this type of active and predictive processing is a part of the puzzle of how sensory systems work in concert with motor behavior to serve the goals of the organism.
Reviewer #3 (Public Review):
This manuscript examines one important and at the same time little investigated question in vision science: what happens to the processing of the foveal input right before the onset of a saccade. This is clearly something of relevance as humans perform saccades about 3 times every second. Whereas what happens to visual perception in the visual periphery at the saccade goal is well characterized, little is known about what happens at the very center of gaze, which represents the future retinal location where the saccade target will be viewed at high resolution upon landing. To address this problem the authors implemented an elegant experiment in which they probed foveal vision at different times before the onset of the saccade by using a target, with the same or different orientation with respect to the stimulus at the saccade goal, embedded in dynamic noise. The authors show that foveal processing of the saccade target is initiated before saccade execution resulting in the visual system being more sensitive to foveal stimuli which features match with those of the stimuli at the saccades goal. According to the authors, this process enables a smooth transition of visual perception before and after the saccade. The experiment is well designed and the results are solid, overall I think this work represents a valuable contribution to the field and its results have important implications. My comments below:
- The change in the overall performance between the baseline condition and when the probe is presented after the saccade target is large, but I wonder if there are other unrelated factors that contribute to this difference, for example, simply presenting the probe after vs before the onset of a peripheral stimulus, or the fact that in the baseline the probe is presented right after a fixation marker, but in the other condition there was a longer time interval between the presentation of the marker and the probe transient. The authors should discuss how these confounding factors have been accounted for.
We thank the reviewer for this helpful comment. We would like to clarify that the probe was never presented right after the fixation dot. In the baseline condition, fixation dot and target were separated by 50 ms, i.e., the duration of one noise image. Since the fixation dot was an order of magnitude smaller than the probe (0.3 vs 3 dva in diameter) and since two large-field visual transients caused by the onset of a new background noise image occurred between fixation dot disappearance and probe appearance, we consider it unlikely that the performance difference was caused by any kind of stimulus interaction such as masking. Nonetheless, we had been puzzled by this difference already when inspecting preliminary results and wondered if it may reflect observers’ temporal expectations about the trial sequence. We therefore explicitly instructed and repeatedly reminded observers that the probe could appear before the peripheral target. Since the difference persisted, we ascribed it to a predictive remapping of attention to the fovea during saccade preparation, as we had stated in the Discussion.
Another contributing factor may be that observers approached the oculomotor and perceptual detection tasks sequentially. In early trial phases, they may have prioritized localizing the target and programming the eye movement. After motor planning had been initiated, resources may have been freed up for the foveal detection task. Since on the majority of probe-present trials, the probe appeared after the saccade target, this strategy would have been mostly adaptive. Crucially, however, observers yielded similar incongruent Hit Rates in the baseline and last pre-saccadic time bin (70% vs 74%). While we observed pronounced enhancement in the last pre-saccadic bin, congruent and incongruent Hit Rates in the baseline bin were virtually identical. We therefore conclude that lower overall performance in the baseline bin did not prevent congruency effects from occurring. Instead, congruency effects started developing only after target appearance. We have added this potential explanation to the Results.
- Somewhat related to point 3, the authors conclude that the effects reported here are the result of saccade preparation/execution, however, a control condition in which the saccade is not performed is missing. This leaves me wondering whether the effect is only present during saccade preparation or if it may also be present to some extent or to its full extent when covert attention is engaged, i.e when subjects perform the same task without making a saccade.
Foveal feedback has, as of now, exclusively been demonstrated during fixation (see references in Introduction and Discussion). In most of these studies, it was suggested that these effects (i.e., the foveal representation of a peripheral stimulus) may reflect the automatic preparation of an eye movement that was simply not executed[11,12,14]. Since foveal feedback has been demonstrated during fixation, and since eye movement preparation may influence foveal processing even when the eyes remain stationary, we considered it likely that congruency effects would emerge during fixation. Nonetheless, we agree with the reviewer that an explicit comparison between saccade preparation and fixation would enrich our data set and allow for stronger conclusions. We therefore collected additional data from seven observers. While all remaining experimental parameters were identical to Experiment 1, observers maintained fixation throughout each trial. We found that pre-saccadic foveal enhancement was more pronounced and emerged earlier than foveal enhancement during fixation. We present these data in the Results section (Figure 5) and have updated the Methods section to incorporate this additional experiment. We have furthermore added a paragraph to the Discussion which addresses potential mechanisms of foveal enhancement during fixation and saccade preparation.
Furthermore, the reviewer’s comment helped us realize that we never stated a crucial part of our motivation explicitly. We now do so in the Introduction:
“Despite the theoretical usefulness of such a mechanism, there are reasons to assume that foveal feedback may break down while an eye movement is prepared to a different visual field location. First and foremost, saccade preparation is accompanied with an obligatory shift of attention to the saccade target[6-8] which in turn has been shown to decrease foveal sensitivity[9]. Moreover, the execution of a rapid eye movement induces brief motion signals on the retina[20] which may mask or in other ways interfere with the pre-saccadic prediction signal. On a more conceptual level, the recruitment of foveal processing as an ‘active blackboard’[21] may become obsolete in the face of an imminent foveation of relevant peripheral stimuli – unless, of course, foveal processing serves the establishment of trans-saccadic visual continuity.”
We believe that the additional data and the revisions to the Introduction and Discussion have strengthened our manuscript and thank the reviewer for this comment.
- Differently from other tasks addressing pre-saccadic perception in the literature here subjects do not have to discriminate the peripheral stimulus at the saccade goal, and most processing resources are presumably focused at the foveal location. Could this have influenced the results reported here?
This is true. We intentionally made the features of the peripheral target as task-irrelevant as possible, contrary to previous investigations. We wanted to ensure that the enhancement we find would be automatic and not induced by a peripheral discrimination task, as we state in the Discussion and the Methods. We agree that the foveal detection task likely focused processing resources on the center of gaze in Experiment 1. In Experiment 2, however, we measured the spatial profile of enhancement which involved two different conditions:
- In each observer’s first six sessions, the probe could be presented anywhere on a horizontal axis of 9 dva length. On a given trial, an observer could not predict where it would appear, and therefore could not strategically allocate their attention. Nonetheless, enhancement of target-congruent orientation information was tuned to the fovea.
- In the final, seventh session, the probe appeared exclusively in one of two possible peripheral locations: 3 dva to the left or 3 dva to the right of the screen center. Observers were explicitly informed that the probe would never appear foveally, and processing resources should therefore have been allocated to the peripheral probe locations. The general performance level in this condition was comparable to performance in the fovea (see reply to the next comment). Nonetheless, we did not find peripheral enhancement of target-congruent information.
Importantly, the magnitude of the foveal congruency effect in the PRE-only condition of Experiment 1 (i.e., when the target disappeared before the eyes landed on it) was comparable to the foveal congruency effect in Experiment 2 (PRE-only throughout), suggesting that the format of the task – i.e., purely foveal detection or foveal and peripheral detection – did not alter our findings.
- The spatial profile of the enhancement is very interesting and it clearly shows that the enhancement is limited to a central region. To which extent this profile is influenced by the fact that the probe was presented at larger eccentricities and therefore was less visible at 4.5 deg than it was at 0 deg? According to the caption, when the probe was presented more eccentrically the performance was raised to a foveal level by adaptively increasing probe transparency. This is not clear, was this done separately based on performance at baseline? Does this mean that the contrast of the stimulus was different for the points at +- 3 dva but the performance was comparable at baseline? Please explain.
Based on the previous comment and comments of Reviewer #2, we realize that we should have explained this condition more extensively in the main text rather than in the Methods and have adapted the manuscript accordingly. As stated in our reply to the previous comment, Experiment 2 involved one session in which we addressed whether the lack of parafoveal/peripheral enhancement could be due to a simple decrease in acuity as mentioned by the reviewer. Observers were explicitly informed that the to-be detected stimulus (the probe) would appear either 3 dva to the left or right but never in the screen center and were shown slowed-down example trials for illustration. Observers then performed a staircase procedure which was targeted at determining the probe contrast at which performance for parafoveal target-incongruent probes would be just as high as foveal performance for target-incongruent probes had been in the previous six sessions. While the foveal probe was presented at a median opacity of 28.3±7.6%, an opacity of 39.0±11.1% was required to achieve the same performance level at a 3 dva eccentricity. Therefore, the gray curve in Figure 2D that represents incongruent Hits reaches its peak just under 80% on the y-axis. The gray dots at ±3 dva also range at ~80% on the y-axis. The performance level for target-incongruent probes (‘baseline’ here) in the parafovea is thus equal to foveal performance for target-incongruent probes. Target-congruent parafoveal feature information had the same “chance” to be enhanced as foveal information in the preceding sessions. Despite an equation of performance, we found no parafoveal enhancement. This suggests that enhancement is a true consequence of visual field location and not simply mediated by visual acuity at that location.
- The enhancement is significant within a region of 6.4 dva around the center of gaze. This is a rather large region, especially considering that it extends also in the direction opposite to the saccade. I was expecting the enhancement to be more confined to the central foveal region. Was the effect shown in Figure 2D influenced by the fact that saccades in this task were characterized by a large undershoot (Fig 1 D)? Did the effect change if only saccades landing closer to the target were included in the analysis? There may not be enough data for resolving the time course, but maybe there are differences in the size of the main effect.
Width of the profile: In general, the width of the enhancement profile is likely to be influenced by two experimental/analysis choices: the size of the probe stimulus presented during the experiment and the width of the moving window combining adjacent probe locations for analysis.
Probe size: Since the probe itself had a comparably large diameter of 3 dva, even the leftmost significant point at -2.6 dva could be explained by an enhancement of the foveal portion of the probe. We had mentioned this briefly in the Discussion but realize that this point is crucial and should be made more explicit. Moving window width: We designed the experiment with the intention to densely sample a range of spatial locations during data collection and combine a certain number of adjacent locations using a moving window during analysis (see preregistration: https://osf.io/6s24m). To ensure the reliability of every data point, the width of this window was chosen based on how many trials were lost during preprocessing. We chose a window width of 7 locations as this ensured that each data point contained at least 30 trials on an individual-observer level. Nonetheless, the width of the resulting enhancement profile depends on the width of the moving window:
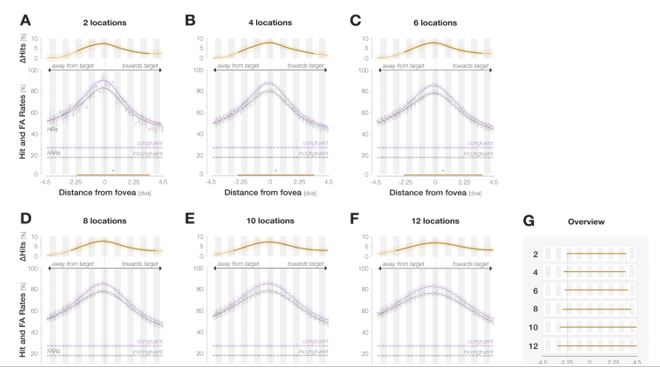
We added these caveats to the Results section and incorporated the figure above into the Supplements. We now state explicitly that…
“the main conclusions that can be drawn are that enhancement i) peaks in the center of gaze, ii) is not uniform throughout the tested spatial range as, for instance, global feature-based attention would predict, and iii) is asymmetrical, extending further towards the saccade target than away from it.”
For the above reasons, the absolute width of the profile should be interpreted with caution.
Saccadic landing accuracy: To address the reviewer’s question, we inspected the spatial enhancement profile separately for trials in which the saccade landed on the target (i.e., within a radius of 1.5 dva from its center) or off-target but still within the accepted landing area. This trial separation criterion, besides appearing meaningful, ensured that all observers contributed trials to every data point. We had never resolved the time course in this experiment and could therefore not collapse across time points as suggested by the reviewer. To increase the number of trials per data point, we instead increased the width of the moving window sliding across locations from 6 to 9 neighboring locations (but see caveat above).
Considering only saccades that landed on the target (‘accurate’; A) yielded significant enhancement from -2.6 to 2.1 dva and from 3.2 dva throughout the measured range towards the saccade target. Saccades that landed off-target (‘inaccurate’; B) showed a more pronounced asymmetry. When only considering inaccurate saccades, enhancement reached significance between -1.1 and 4.4 dva.
The increased asymmetry for inaccurate saccades may be related to predictive remapping: since inaccurate saccades were hypometric on average, the predictively remapped location of the target was shifted towards the target by the magnitude of the undershoot. Asymmetric enhancement would therefore have boosted congruency at the remapped target location across all trials. In consequence, we inspected if aligning probe locations to the remapped target location on an individual-trial level would lead to a narrower profile for inaccurate saccades. This was not the case. Instead, we observed two parafoveal maxima (C). Their position on the x-axis equals the mean remapping-dependent leftwards (2.0 dva) and rightwards (1.9 dva) displacement across trials. In other words, they correspond to the pre-saccadic center of gaze. Note that these profiles could not be fitted with a mixture of Gaussians and were fitted using polynomials instead.
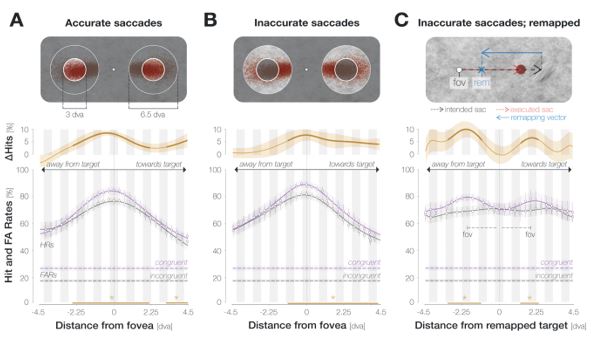
In sum, while we do not observe a clear narrowing of the enhancement profile for accurate saccades, the profile’s asymmetry is more pronounced for inaccurate eye movements. An increase in asymmetry could bear functional advantages since it would boost congruency at the remapped target location across all trials. Importantly though, this adjustment seems to rely on an estimate of average rather than single-trial saccade characteristics: aligning probe locations to the remapped attentional locus on an individual trial level provides further evidence that, irrespective of individual saccade endpoints, enhancement was aligned to the fovea. We have added these analyses to the Results section (Figure 3). We have also added the remapped profiles for all saccades and accurate saccades only to the Supplements.
- Is the size of the enhanced region around the center of gaze related to the precision of saccades? Presumably, if saccades are less precise a larger enhanced area may be more beneficial.
This is a very interesting point. To address this question, we estimated each observer’s saccadic precision by computing bivariate kernel densities from their saccade landing coordinates. As we measured the horizontal extent of enhancement in our experiment, we defined the horizontal bandwidth as an estimate of saccadic imprecision. To estimate the size of the enhanced region for each observer, we created 10,000 bootstrapping samples for each observer’s congruent and incongruent HRs (4 locations combined at each step) We then determined the difference between the bootstrapped congruent and incongruent HRs and defined significantly enhanced locations as all locations for which <= 5% of these differences fell below zero. We then defined the width of the enhancement profile as the maximum number of consecutive significant locations.
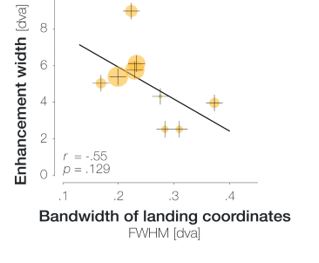
Instead of a positive correlation, we observed a negative correlation between the bandwidth of landing coordinates (i.e., saccadic imprecision) and the size of the enhanced window (r = -.56, p = .117). In other words, there was a non-significant tendency that the less precise an observer’s saccades, the narrower their estimated region of enhancement. We furthermore inspected the magnitude of enhancement per position within in the enhanced region. To do so, we computed the mean difference between congruent and incongruent HR across all positions in the enhanced region. The sizes of the orange circles in the figure above represent the resulting values (ranging from 2.9% to 13.3%). As saccadic precision decreases, the magnitude of enhancement per data point in the enhanced region tends to decrease as well. We therefore suggest that high saccadic precision is a sign of efficient oculomotor programming, which in turn allows peri-saccadic perceptual processes to operate more effectively. We added this analysis to the Supplements and refer to it in the Results section of the revised manuscript.
-
Evaluation Summary:
This study measures the strength, timing, feature specificity and for the first time - spatial extent - of pre-saccadic processing of peripheral target information at the fovea, when fixation is not constrained. The authors conclude that saccade preparation causes feature-specific pre-saccadic visual enhancement restricted largely to the center of gaze. The manuscript is based on a rigorously curated dataset, it is well written and illustrated, and will be of potential interest to a broad readership of vision scientists, neuroscientists, and modelers. However, some key weaknesses in the interpretation of data were identified, which call into question the conclusion that the foveal enhancement observed can only be explained in terms of saccade preparation, thus weakening our ability to infer its mechanistic substrates.
Evaluation Summary:
This study measures the strength, timing, feature specificity and for the first time - spatial extent - of pre-saccadic processing of peripheral target information at the fovea, when fixation is not constrained. The authors conclude that saccade preparation causes feature-specific pre-saccadic visual enhancement restricted largely to the center of gaze. The manuscript is based on a rigorously curated dataset, it is well written and illustrated, and will be of potential interest to a broad readership of vision scientists, neuroscientists, and modelers. However, some key weaknesses in the interpretation of data were identified, which call into question the conclusion that the foveal enhancement observed can only be explained in terms of saccade preparation, thus weakening our ability to infer its mechanistic substrates.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. The reviewers remained anonymous to the authors.)
-
Reviewer #1 (Public Review):
In their paper, Kroell and Rolfs use a set of sophisticated psychophysical experiments in visually-intact observers, to show that visual processing at the fovea within the 250ms or so before saccading to a peripheral target containing orientation information, is influenced by orientation signals at the target. Their approach straddles the boundary between enforcing fixation throughout stimulus presentation (a standard in the field) and leaving it totally unconstrained. As such, they move the field of saccade pre-processing towards active vision in order to answer key questions about whether the fovea predicts features at the gaze target, over what time frame, with what precision, and over what spatial extent around the foveal center. The results support the notion that there is feature-selective enhancement …
Reviewer #1 (Public Review):
In their paper, Kroell and Rolfs use a set of sophisticated psychophysical experiments in visually-intact observers, to show that visual processing at the fovea within the 250ms or so before saccading to a peripheral target containing orientation information, is influenced by orientation signals at the target. Their approach straddles the boundary between enforcing fixation throughout stimulus presentation (a standard in the field) and leaving it totally unconstrained. As such, they move the field of saccade pre-processing towards active vision in order to answer key questions about whether the fovea predicts features at the gaze target, over what time frame, with what precision, and over what spatial extent around the foveal center. The results support the notion that there is feature-selective enhancement centered on the center of gaze, rather than on the predictively remapped location of the target. The results further show that this enhancement extends about 3 deg radially from the foveal center and that it starts ~ 200ms or so before saccade onset. They also show that this enhancement is reinforced if the target remains present throughout the saccade. The hypothesized implications of these findings are that they could enable continuity of perception trans-saccadically and potentially, improve post-saccadic gaze correction.
Strengths:
The findings appear solid and backed up by converging evidence from several experimental manipulations. These included several approaches to overcome current methodological constraints to the critical examination of foveal processing while being careful not to interfere with saccade planning and performance. The authors examined the spatial frequency characteristics of the foveal enhancement relative, hit rates and false alarm rates for detecting a foveal probe that was congruent or incongruent in terms of orientation to the peripheral saccade target embedded in flickering, dynamic noise (i/f )images. While hit rates are relatively easy to interpret, the authors also reconstructed key features of the background noise to interpret false alarms as reflecting foveal enhancement that could be correlated with target orientation signals. The study also - in an extensive Supplementary Materials section - uses appropriate statistical analyses and controls for multiple factors impacting experimental/stimulus design and analysis. The approach, as well as the level of care towards experimental details provided in this manuscript, should prove welcome and useful for any other investigators interested in the questions posed.Weaknesses:
I find no major weaknesses in the experiments, analyses or interpretations. The conclusions of the paper appear well supported by the data. My main suggestion would be to see a clearer discussion of the implications of the present findings for truly naturalistic, visually-guided performance and action. Please consider the implication of the phenomena and behaviors reported here when what is located at the gaze center (while peripheral targets are present), is not a noisy, relatively feature-poor, low-saliency background, but another high-saliency target, likely crowded by other nearby targets. As such, a key question that emerges and should be addressed in the Discussion at least is whether the fovea's role described in the present experiments is restricted to visual scenarios used here, or whether they generalize to the rather different visual environments of everyday life. -
Reviewer #2 (Public Review):
Human and primates move their eyes with rapid saccades to reposition the high-resolution region of the retina, the fovea, over objects of interest. Thus, each saccade involves moving the fovea from a pre-saccadic location to a saccade target. Although it has been long known that saccades profoundly alter visual processing at the time of saccade, scientists simply do not know how the brain combines information across saccades to support our normal perceptual experience. This paper addresses a piece of that puzzle by examining how eye movements affect processing at the fovea before it moves. Using a dynamic noise background and a dual psychophysical task, the authors probe both the performance and selectivity of visual processing for orientation at the fovea in the few hundred milliseconds preceding a saccade. …
Reviewer #2 (Public Review):
Human and primates move their eyes with rapid saccades to reposition the high-resolution region of the retina, the fovea, over objects of interest. Thus, each saccade involves moving the fovea from a pre-saccadic location to a saccade target. Although it has been long known that saccades profoundly alter visual processing at the time of saccade, scientists simply do not know how the brain combines information across saccades to support our normal perceptual experience. This paper addresses a piece of that puzzle by examining how eye movements affect processing at the fovea before it moves. Using a dynamic noise background and a dual psychophysical task, the authors probe both the performance and selectivity of visual processing for orientation at the fovea in the few hundred milliseconds preceding a saccade. They find that hit rates and false alarm rates are dynamically and automatically modulated by the saccade planning. By taking advantage of the specific sequence of noise shown on each trial, they demonstrate that the tuning of foveal processing is affected by the orientation of the saccade target suggesting foveal specific feedback.
A major strength of the paper is the experimental design. The use of dynamic filtered noise to probe perceptual processing is a clever way of measuring the dynamics of selectivity at the fovea during saccade preparation. The use of a dual-task allows the authors to evaluate the tuning of foveal processing as well and how it depends on the peripheral target orientation. They show compellingly that the orientation of the saccade target (the future location of the fovea) affects processing at the fovea before it moves.
There are two weaknesses with the paper in its current form. The first is that the key claim of foveal "enhancement" relies on the tuning of the false alarms. A more standard measure of enhancement would be to look at the sensitivity, or d-prime, of the performance on the task. In this study, hits and false alarms increase together, which is traditionally interpreted as a criterion shift and not an enhancement. However, because of the external noise, false alarms are driven by real signals. The authors are aware of this and argue that the fact that the false alarms are tuned indicates enhancement. But it is unclear to me that a criterion shift wouldn't also explain this tuning and the change in the noise images. For example, in a task with 4 alternative choices (Present/Congruent, Present/Incongruent, Absent/Congruent, Absent/Incongruent), shifting the criterion towards the congruent target would increase hits and false alarms for that target and still result in a tuned template (because that template is presumably what drove the decision variable that the adjusted criterion operates on). I believe this weakness could be addressed with a computational model that shows that a criterion shift on the output of a tuned template cannot produce the pattern of hits and false alarms.
The second weakness is that the author's claim that feedback is spatially selective to the fovea is confounded by the fact that acuity and contrast sensitivity are higher in the fovea. Therefore, the subject's performance would already be spatially tuned. Even the very central degree, the foveola, is inhomogeneous. Thus, finding spatially-tuned sensitivity to the probes may simply indicate global feature gain on top of already spatially tuned processing in the fovea. Another possible explanation that is consistent with the "no enhancement" interpretation is that the fovea has increased. This is consistent with the observation that the congruency effects were aligned to the center of gaze and not the saccade endpoint. It looks from the Gaussian fits that a single gain parameter would explain the difference in the shape of the congruent and incongruent hit rates, but I could not figure out if this was explicitly tested from the existing methods. Additional experiments without prepared saccades would be an easy way to address this issue. Is the hit rate tuned when there is no saccade preparation? If so, it seems likely that the spatial selectivity is not tuned feedback, but inhomogeneous feedforward processing.
This paper is important because it compellingly demonstrates that visual processing in the fovea anticipates what is coming once the eyes move. The exact form of the modulation remains unclear and the authors could do more to support their interpretations. However, understanding this type of active and predictive processing is a part of the puzzle of how sensory systems work in concert with motor behavior to serve the goals of the organism.
-
Reviewer #3 (Public Review):
This manuscript examines one important and at the same time little investigated question in vision science: what happens to the processing of the foveal input right before the onset of a saccade. This is clearly something of relevance as humans perform saccades about 3 times every second. Whereas what happens to visual perception in the visual periphery at the saccade goal is well characterized, little is known about what happens at the very center of gaze, which represents the future retinal location where the saccade target will be viewed at high resolution upon landing. To address this problem the authors implemented an elegant experiment in which they probed foveal vision at different times before the onset of the saccade by using a target, with the same or different orientation with respect to the …
Reviewer #3 (Public Review):
This manuscript examines one important and at the same time little investigated question in vision science: what happens to the processing of the foveal input right before the onset of a saccade. This is clearly something of relevance as humans perform saccades about 3 times every second. Whereas what happens to visual perception in the visual periphery at the saccade goal is well characterized, little is known about what happens at the very center of gaze, which represents the future retinal location where the saccade target will be viewed at high resolution upon landing. To address this problem the authors implemented an elegant experiment in which they probed foveal vision at different times before the onset of the saccade by using a target, with the same or different orientation with respect to the stimulus at the saccade goal, embedded in dynamic noise. The authors show that foveal processing of the saccade target is initiated before saccade execution resulting in the visual system being more sensitive to foveal stimuli which features match with those of the stimuli at the saccades goal. According to the authors, this process enables a smooth transition of visual perception before and after the saccade. The experiment is well designed and the results are solid, overall I think this work represents a valuable contribution to the field and its results have important implications. My comments below:
1. The change in the overall performance between the baseline condition and when the probe is presented after the saccade target is large, but I wonder if there are other unrelated factors that contribute to this difference, for example, simply presenting the probe after vs before the onset of a peripheral stimulus, or the fact that in the baseline the probe is presented right after a fixation marker, but in the other condition there was a longer time interval between the presentation of the marker and the probe transient. The authors should discuss how these confounding factors have been accounted for.
2. Somewhat related to point 3, the authors conclude that the effects reported here are the result of saccade preparation/execution, however, a control condition in which the saccade is not performed is missing. This leaves me wondering whether the effect is only present during saccade preparation or if it may also be present to some extent or to its full extent when covert attention is engaged, i.e when subjects perform the same task without making a saccade.
3. Differently from other tasks addressing pre-saccadic perception in the literature here subjects do not have to discriminate the peripheral stimulus at the saccade goal, and most processing resources are presumably focused at the foveal location. Could this have influenced the results reported here?
4. The spatial profile of the enhancement is very interesting and it clearly shows that the enhancement is limited to a central region. To which extent this profile is influenced by the fact that the probe was presented at larger eccentricities and therefore was less visible at 4.5 deg than it was at 0 deg? According to the caption, when the probe was presented more eccentrically the performance was raised to a foveal level by adaptively increasing probe transparency. This is not clear, was this done separately based on performance at baseline? Does this mean that the contrast of the stimulus was different for the points at +- 3 dva but the performance was comparable at baseline? Please explain.
5. The enhancement is significant within a region of 6.4 dva around the center of gaze. This is a rather large region, especially considering that it extends also in the direction opposite to the saccade. I was expecting the enhancement to be more confined to the central foveal region. Was the effect shown in Figure 2D influenced by the fact that saccades in this task were characterized by a large undershoot (Fig 1 D)? Did the effect change if only saccades landing closer to the target were included in the analysis? There may not be enough data for resolving the time course, but maybe there are differences in the size of the main effect.
6. Is the size of the enhanced region around the center of gaze related to the precision of saccades? Presumably, if saccades are less precise a larger enhanced area may be more beneficial.
-
