Cognitive experience alters cortical involvement in goal-directed navigation
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
In this study, the role of different cortical areas on three distinct tasks all relying on the same virtual maze set-up was examined using optogenetic interventions and calcium imaging. The paper is potentially of interest to people interested in understanding the neural substrates of learning and how these can be impacted by previous knowledge and experience of stimuli. It could also be of use to behavioral neuroscientists when considering possible order effects of experiments.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Neural activity in the mammalian cortex has been studied extensively during decision tasks, and recent work aims to identify under what conditions cortex is actually necessary for these tasks. We discovered that mice with distinct cognitive experiences, beyond sensory and motor learning, use different cortical areas and neural activity patterns to solve the same navigation decision task, revealing past learning as a critical determinant of whether cortex is necessary for goal-directed navigation. We used optogenetics and calcium imaging to study the necessity and neural activity of multiple cortical areas in mice with different training histories. Posterior parietal cortex and retrosplenial cortex were mostly dispensable for accurate performance of a simple navigation task. In contrast, these areas were essential for the same simple task when mice were previously trained on complex tasks with delay periods or association switches. Multiarea calcium imaging showed that, in mice with complex-task experience, single-neuron activity had higher selectivity and neuron–neuron correlations were weaker, leading to codes with higher task information. Therefore, past experience is a key factor in determining whether cortical areas have a causal role in goal-directed navigation.
Article activity feed
-

Author Response
Reviewer #1 (Public Review):
The role of the parietal (PPC), the retrospenial (RSP) and the the visual cortex (S1) was assessed in three tasks corresponding a simple visual discrimination task, a working-memory task and a two-armed bandit task all based on the same sensory-motor requirements within a virtual reality framework. A differential involvement of these areas was reported in these tasks based on the effect of optogenetic manipulations. Photoinhibition of PPC and RSP was more detrimental than photoinhibition of S1 and more drastic effects were observed in presumably more complex tasks (i.e. working-memory and bandit task). If mice were trained with these more complex tasks prior to training in the simple discrimination task, then the same manipulations produced large deficits suggesting that switching from one …
Author Response
Reviewer #1 (Public Review):
The role of the parietal (PPC), the retrospenial (RSP) and the the visual cortex (S1) was assessed in three tasks corresponding a simple visual discrimination task, a working-memory task and a two-armed bandit task all based on the same sensory-motor requirements within a virtual reality framework. A differential involvement of these areas was reported in these tasks based on the effect of optogenetic manipulations. Photoinhibition of PPC and RSP was more detrimental than photoinhibition of S1 and more drastic effects were observed in presumably more complex tasks (i.e. working-memory and bandit task). If mice were trained with these more complex tasks prior to training in the simple discrimination task, then the same manipulations produced large deficits suggesting that switching from one task to the other was more challenging, resulting in the involvement of possibly larger neural circuits, especially at the cortical level. Calcium imaging also supported this view with differential signaling in these cortical areas depending on the task considered and the order to which they were presented to the animals. Overall the study is interesting and the fact that all tasks were assessed relying on the same sensory-motor requirements is a plus, but the theoretical foundations of the study seems a bit loose, opening the way to alternate ways of interpreting the data than "training history".
- Theoretical framework:
The three tasks used by the authors should be better described at the theoretical level. While the simple task can indeed be considered a visual discrimination task, the other two tasks operationally correspond to a working-memory task (i.e. delay condition which is indeed typically assessed in a Y- or a T-maze in rodent) or a two-armed bandit task (i.e. the switching task), respectively. So these three tasks are qualitatively different, are therefore reliant on at least partially dissociable neural circuits and this should be clearly analyzed to explain the rationale of the focus on the three cortical regions of interest.
We are glad to see that the reviewer finds our study interesting overall and sees value in the experimental design. We agree that in the previous version, we did not provide enough motivation for the specific tasks we employed and the cortical areas studied.
Navigating to reward locations based on sensory cues is a behavior that is crucial for survival and amenable to a head-fixed laboratory setting in virtual reality for mice. In this context of goal-directed navigation based on sensory cues, we chose to center our study on posterior cortical association areas, PPC and RSC, for several reasons. RSC has been shown to be crucial for navigation across species, poised to enable the transformation between egocentric and allocentric reference frames and to support spatial memory across various timescales (Alexander & Nitz, 2015; Fischer et al., 2020; Pothuizen et al., 2009; Powell et al., 2017). It furthermore has been shown to be involved in cognitive processes beyond spatial navigation, such as temporal learning and value coding (Hattori et al., 2019; Todd et al., 2015), and is emerging as a crucial region for the flexible integration of sensory and internal signals (Stacho & ManahanVaughan, 2022). It thus is a prime candidate area in the study of how cognitive experience may affect cortical involvement in goal-directed navigation.
RSC is heavily interconnected with PPC, which is generally thought to convert sensory cues into actions (Freedman & Ibos, 2018) and has been shown to be important for navigation-based decision tasks (Harvey et al., 2012; Pinto et al., 2019). Specific task components involving short-term memory have been suggested to cause PPC to be necessary for a given task (Lyamzin & Benucci, 2019), so we chose such task components in our complex tasks to maximize the likelihood of large PPC involvement to compare the simple task to.
One such task component is a delay period between cue and the ultimate choice report, which is a common design in decision tasks (Goard et al., 2016; Harvey et al., 2012; Katz et al., 2016; Pinto et al., 2019). We agree with the reviewer that traditionally such a task would be referred to as a workingmemory task. However, we refrain from using this terminology because it may cause readers to expect that to solve the task, mice use a working-memory dependent strategy in its strictest and most traditional sense, that is mice show no overt behaviors indicative of the ultimate choice until the end of the delay period. If the ultimate choice is apparent earlier, mice may use what is sometimes referred to as an embodiment-based strategy, which by some readers may be seen as precluding working memory. Indeed, in new choice-decoding analyses from the mice’s running patterns, we show that mice start running towards the side of the ultimate choice during the cue period already (Figure 1—figure supplement 1). Regardless of these seemingly early choices, however, we crucially have found much larger performance decrements from inhibition in mice performing the delay task compared to mice performing the simple task, along with lower overall task performance in the delay task, indicating that the insertion of a delay period increased subjective task difficulty. As traditional working-memory versus embodiment-based strategies are not the focus of our study here and do not seem to inform the performance decrements from inhibition, we chose to label the task descriptively with the crucial task parameter rather than with the supposedly underlying cognitive process.
For the switching task, we appreciate that the reviewer sees similarities to a two-armed bandit task. However, in a two-armed bandit task, rewards are typically delivered probabilistically, whereas in our task, cue and action values are constant within each of the two rule blocks, and only the rule, i.e. the cuechoice association, reverses across blocks. This is a crucial distinction because in our design, blocks of Rule A in the switching task are identical to the simple task, with fixed cue-choice associations and guaranteed reward delivery if the correct choice is made, allowing a fair comparison of cortical involvement across tasks.
We have now heavily revised the introduction, results, and discussion sections of the manuscript to better explain the motivation for the tasks and the investigated brain areas. These revisions cover all the points mentioned in this response.
Furthermore, we agree with the reviewer that the three tasks are qualitatively different and likely depend on at least partially dissociable circuits. We consider the large differences in cortical inhibition effects between the simple and the complex tasks as evidence for this notion. We also want to highlight that in fact, we performed task-specific optogenetic manipulations presented in the Supplementary Material to further understand the involvement of different areas in task-specific processes. In what is now Figure 1—figure supplement 4, we restricted inhibition in the delay task to either the cue period only or delay period only, finding that interestingly, PPC or RSC inhibition during either period caused larger performance drops than observed in the simple task. We also performed epoch-specific inhibition of PPC in the switching task, targeting specifically reward and inter-trial-interval periods following rule switches, in what is now Figure 1—figure supplement 5. With such PPC inhibition during the ITI, we observed no effect on performance recovery after rule switches and thus found PPC activity to be dispensable for rule updates.
For the working-memory task we do not know the duration of the delay but this really is critical information; per definition, performance in such a task is delay-dependent, this is not explored in the paper.
We thank the reviewer for pointing out the lack of information on delay duration and have now added this to the Methods section.
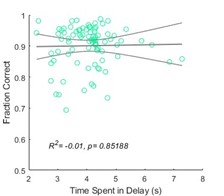
We agree that in classical working memory tasks where the delay duration is purely defined by the experimenter and varied throughout a session, performance is typically dependent on delay duration. However, in our delay task, the delay distance is kept constant, and thus the delay is not varied by the experimenter. Instead, the time spent in the delay period is determined by the mouse, and the only source of variability in the time spent in the delay period is minor differences in the mice’s running speeds across trials or sessions. Notably, the differences in time in the delay period were greatest between mice because some mice ran faster than others. Within a mouse, the time spent in the delay period was generally rather consistent due to relatively constant running speeds. Also, because the mouse had full control over the delay duration, it could very well speed up its running if it started to forget the cue and run more slowly if it was confident in its memory. Thus, because the delay duration was set by the mouse and not the experimenter, it is very challenging or impossible to interpret the meaning and impact of variations in the delay duration. Accordingly, we had no a priori reason to expect a relationship between task performance and delay duration once mice have become experts at the delay task. Indeed, we do not see such a relationship in our data (see plot here, n = 85 sessions across 7 mice). In order to test the effect of delay duration on behavioral performance, we would have to systematically change the length of the delay period in the maze, which we did not do and which would require an entirely new set of experiments.
Also, the authors heavily rely on "decision-making" but I am genuinely wondering if this is at all needed to account for the behavior exhibited by mice in these tasks (it would be more accurate for the bandit task) as with the perspective developed by the authors, any task implies a "decision-making" component, so that alone is not very informative on the nature of the cognitive operations that mice must compute to solve the tasks. I think a more accurate terminology in line with the specific task considered should be employed to clarify this.
We acknowledge that the previous emphasis on decision-making may have created expectations that we demonstrate effects that are specific to the ‘decision-making’ aspect of a decision task. As we do not isolate the decision-making process specifically, we have substantially revised our wording around the tasks and removed the emphasis on decision-making, including in the title. Rather than decision-making, we now highlight the navigational aspect of the tasks employed.
The "switching"/bandit task is particularly interesting. But because the authors only consider trials with highest accuracy, I think they are missing a critical component of this task which is the balance between exploiting current knowledge and the necessity to explore alternate options when the former strategy is no longer effective. So trials with poor performance are thus providing an essential feedback which is a major drive to support exploratory actions and a critical asset of the bandit task. There is an ample literature documenting how these tasks assess the exploration/exploitation trade-off.
We completely agree with the reviewer that the periods following rule switches are an essential part of the switching task and of high interest. Indeed, ongoing work in the lab is carefully quantifying the mice’s strategy in this task and exploring how mice use errors after switches to update their belief about the rule. In this project, however, a detailed quantification of switching task strategy seemed beyond the scope because our focus was on training history and not on the specifics of each task. While we agree with the reviewer about the interesting nature of the switching period, it would be too much for a single paper to investigate the detailed mechanisms of each task on top of what we already report for training history. Instead, we have now added quantifications of performance recovery after rule switches in Figure 1— figure supplement 2, showing that rule switches cause below-chance performance initially, followed by recovery within tens of trials.
- Training history vs learning sets vs behavioral flexibility:
The authors consider "training history" as the unique angle to interpret the data. Because the experimental setup is the same throughout all experiments, I am wondering if animals are just simply provided with a cognitive challenge assessing behavioral flexibility given that they must identify the new rule while restraining from responding using previously established strategies. According to this view, it may be expected for cortical lesions to be more detrimental because multiple cognitive processes are now at play.
It is also possible that animals form learning sets during successive learning episodes which may interfere with or facilitate subsequent learning. Little information is provided regarding learning dynamics in each task (e.g. trials to criterion depending on the number of tasks already presented) to have a clear view on that.
We thank the reviewer for raising these interesting ideas. We have now evaluated these ideas in the context of our experimental design and results. One of the main points to consider is that for mice transitioned from either of the complex tasks to the simple task, the simple task is not a novel task, but rather a well-known simplification of the previous tasks. Mice that are experts on the delay task have experienced the simple task, i.e. trials without a delay period, during their training procedure before being exposed to delay periods. Switching task expert mice know the simple task as one rule of the switching task and have performed according to this rule in each session prior to the task transition. Accordingly, upon to the transition to the simple task, both delay task expert mice and switching task expert mice perform at very high levels on the very first simple task session. We now quantify and report this in Figure 2—figure supplement 1 (A, B). This is crucial to keep in mind when assessing ‘learning sets’ or ‘behavioral flexibility’ as possible explanations for the persistent cortical involvement after the task transitions. In classical learning sets paradigms, animals are exposed to a series of novel associations, and the learning of previous associations speeds up the learning of subsequent ones (Caglayan et al., 2021; Eichenbaum et al., 1986; Harlow, 1949). This is a distinct paradigm from ours because the simple task does not contain novel associations that are new to the mice already trained on the complex tasks. Relatedly, the simple task is unlikely to present a challenge of behavioral flexibility to these mice given our experimental design and the observation of high simple task performance in the first session after the task transition.
We now clarify these points in the introduction, results, and discussion sections, also acknowledging that it will be of interest for future work to investigate how learning sets may affect cortical task involvement.
- Calcium imaging data versus interventions:
The value of the calcium imaging data is not entirely clear. Does this approach bring a new point to consider to interpret or conclude on behavioral data or is it to be considered convergent with the optogenetic interventions? Very specific portions of behavioral data are considered for these analyses (e.g. only highly successful trials for the switching/bandit task) and one may wonder if considering larger or different samples would bring similar insights. The whole take on noise correlation is difficult to apprehend because of the same possible interpretation issue, does this really reflect training history, or that a new rule now must be implemented or something else? I don't really get how this correlative approach can help to address this issue.
We thank the reviewer for pointing out that the relationship between the inhibition dataset and calcium imaging dataset is not clear enough. We restricted analyses of inhibition and calcium imaging data in the switching task to the identical cue-choice associations as present in the simple task (i.e. Rule A trials of the switching task). We did this because we sought to make the fairest and most convincing comparison across tasks for both datasets. However, we can now see that not reporting results with trials from the other rule causes concerns that the reported differences across tasks may only hold for a specific subset of trials.
We have now added analyses of optogenetic inhibition effects and calcium imaging results considering Rule B trials. In Figure 1—figure supplement 2, we show that when considering only Rule B trials in the switching task, effects of RSC or PPC inhibition on task performance are still increased relative to the ones observed in mice trained on and performing the simple task. We also show that overall task performance is lower in Rule B trials of the switching task than in the simple task, mirroring the differences across tasks when considering Rule A trials only.
We extended the equivalent comparisons to the calcium imaging dataset, only considering Rule B trials of the switching task in Figure 4—figure supplement 3. With Rule B trials only, we still find larger mean activity and trial-type selectivity levels in RSC and PPC, but not in V1, compared to the simple task, as well as lower noise correlations. We thus find that our conclusions about area necessity and activity differences across tasks hold for Rule B trials and are not due to only considering a subset of the switching task data.
In Figure 4—figure supplement 4, we further leverage the inclusion of Rule B trials and present new analyses of different single-neuron selectivity categories across rules in the switching task, reporting a prevalence of mixed selectivity in our dataset.
Furthermore, to clarify the link between the optogenetic inhibition and the calcium imaging datasets, we have revised the motivation for the imaging dataset, as well as the presentation of its results and discussion. Investigating an area’s neural activity patterns is a crucial first step towards understanding how differential necessity of an area across tasks or experience can be explained mechanistically on a circuit level. We now elaborate on the fact that mechanistically, changes in an area’s necessity may or may not be accompanied by changes in activity within that area, as previous work in related experimental paradigms has reported differences in necessity in the absence of differences in activity (Chowdhury & DeAngelis, 2008; Liu & Pack, 2017). This phenomenon can be explained by differences in the readout of an area’s activity. We now make more explicit that in contrast to the scenario where only the readout changes, we find an intriguing correspondence between increased necessity (as seen in the inhibition experiments) and increased activity and selectivity levels (as seen in the imaging experiments) in cortical association areas depending on the current task and previous experience. Rather than attributing the increase in necessity solely to these observed changes in activity, we highlight that in the simple task condition already, cortical areas contain a high amount of task information, ruling out the idea that insufficient local information would cause the small performance deficits from inhibition. Our results thus suggest that differential necessity across tasks and experience may still require changes at the readout level despite changes in local activity. We view our imaging results as an exciting first step towards a mechanistic understanding of how cognitive experience affects cortical necessity, but we stress that future work will need to test directly the relationship between cortical necessity and various specific features of the neural code.
Reviewer #2 (Public Review):
The authors use a combination of optogenetics and calcium imaging to assess the contribution of cortical areas (posterior parietal cortex, retrosplenial cortex, S1/V1) on a visual-place discrimination task. Headfixed mice were trained on a simple version of the task where they were required to turn left or right depending on the visual cue that was present (e.g. X = go left; Y = go right). In a more complex version of the task the configurations were either switched during training or the stimuli were only presented at the beginning of the trial (delay).
The authors found that inhibiting the posterior parietal cortex and retrosplenial cortex affected performance, particularly on the complex tasks. However, previous training on the complex tasks resulted in more pronounced impairments on the simple task than when behaviourally naïve animals were trained/tested on a simple task. This suggests that the more complex tasks recruit these cortical areas to a greater degree, potentially due to increased attention required during the tasks. When animals then perform the simple version of the task their previous experience of the complex tasks is transferred to the simple task resulting in a different pattern of impairments compared to that found in behaviorally naïve animals.
The calcium imaging data showed a similar pattern of findings to the optogenetic study. There was overall increased activity in the switching tasks compared to the simple tasks consistent with the greater task demands. There was also greater trial-type selectivity in the switching task compared to the simple task. This increased trial-type selectivity in the switching tasks was subsequently carried forward to the simple task so that activity patterns were different when animals performed the simple task after experiencing the complex task compared to when they were trained on the simple task alone
Strengths:
The use of optogenetics and calcium-imaging enables the authors to look at the requirement of these brain structures both in terms of necessity for the task when disrupted as well as their contribution when intact.
The use of the same experimental set up and stimuli can provide a nice comparison across tasks and trials.
The study nicely shows that the contribution of cortical regions varies with task demands and that longerterm changes in neuronal responses c can transfer across tasks.
The study highlights the importance of considering previous experience and exposure when understanding behavioural data and the contribution of different regions.
The authors include a number of important controls that help with the interpretation of the findings.
We thank the reviewer for pointing out these strengths in our work and for finding our main conclusions supported.
Weaknesses:
There are some experimental details that need to be clarified to help with understanding the paper in terms of behavior and the areas under investigation.
The use of the same stimuli throughout is beneficial as it allows direct comparisons with animals experiencing the same visual cues. However, it does limit the extent to which you can extrapolate the findings. It is perhaps unsurprising to find that learning about specific visual cues affects subsequent learning and use of those specific cues. What would be interesting to know is how much of what is being shown is cue specific learning or whether it reflects something more general, for example schema learning which could be generalised to other learning situations. If animals were then trained on a different discrimination with different stimuli would this previous training modify behavior and neural activity in that instance. This would perhaps be more reflective of the types of typical laboratory experiments where you may find an impairment on a more complex task and then go on to rule out more simple discrimination impairments. However, this would typically be done with slightly different stimuli so you don't introduce transfer effects.
We agree with the reviewer that investigating the effects of schema learning on cortical task involvement is an exciting future direction and have now explicitly mentioned this in the Discussion section. As the reviewer points out, however, our study was not designed to test this idea specifically. Because investigating schema learning would require developing and implementing an entirely new set of behavioral task variants, we feel this is beyond the scope of the current work. As to the question of how generalized the effects of cognitive experience are, our data in the run-to-target task suggest that if task settings are sufficiently distinct, cortical involvement can be similarly low regardless of complex task experience (now Figure 3—figure supplement 1). This finding is in line with recent work from (Pinto et al., 2019), where cortical involvement appears to change rapidly depending on major differences in task demands. However, work in MT has shown that previous motion discrimination training using dots can alter MT involvement in motion discrimination of gratings (Liu & Pack, 2017), highlighting that cortical involvement need not be tightly linked to the sensory cue identity.
It is not clear whether length of training has been taken into account for the calcium imaging study given the slow development of neural representations when animals acquire spatial tasks.
We apologize that the training duration and the temporal relationship between task acquisition and calcium imaging was not documented for the calcium imaging dataset. Please see our detailed reply below the ‘recommendations for the authors’ from Reviewer 2 below.
The authors are presenting the study in terms of decision-making, however, it is unclear from the data as presented whether the findings specifically relate to decision making. I'm not sure the authors are demonstrating differential effects at specific decision points.
We understand that the previous emphasis on decision-making may have created expectations that we demonstrate effects that are specific to the ‘decision-making’ aspect of a decision task. As we do not isolate the decision-making process specifically, we have substantially revised our wording around the tasks and removed the emphasis on decision-making, including in the title. Rather than decision-making, we now highlight the navigational aspect of the tasks employed.
While we removed the emphasis on the decision-making process in our tasks, we found the reviewer’s suggestion to measure ‘decision points’ a useful additional behavioral characterization across tasks. So, we quantified how soon a mouse’s ultimate choice can be decoded from its running pattern as it progresses through the maze towards the Y-intersection. We now show these results in Figure 1—figure supplement 1. Interestingly, we found that in the delay task, choice decoding accuracy was already very high during the cue period before the onset of the delay. Nevertheless, we had shown that overall task performance and performance with inhibition were lower in the delay task compared to the simple task. Also, in segment-specific inhibition experiments, we had found that inhibition during only the delay period or only the cue period decreased task performance substantially more than in the simple task, thus finding an interesting absence of differential inhibition effects around decision points. Overall, how early a mouse made its ultimate decision did not appear predictive of the inhibition-induced task decrements, which we also directly quantify in Figure 1—figure supplement 1.
-
Evaluation Summary:
In this study, the role of different cortical areas on three distinct tasks all relying on the same virtual maze set-up was examined using optogenetic interventions and calcium imaging. The paper is potentially of interest to people interested in understanding the neural substrates of learning and how these can be impacted by previous knowledge and experience of stimuli. It could also be of use to behavioral neuroscientists when considering possible order effects of experiments.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
The role of the parietal (PPC), the retrospenial (RSP) and the the visual cortex (S1) was assessed in three tasks corresponding a simple visual discrimination task, a working-memory task and a two-armed bandit task all based on the same sensory-motor requirements within a virtual reality framework. A differential involvement of these areas was reported in these tasks based on the effect of optogenetic manipulations. Photoinhibition of PPC and RSP was more detrimental than photoinhibition of S1 and more drastic effects were observed in presumably more complex tasks (i.e. working-memory and bandit task). If mice were trained with these more complex tasks prior to training in the simple discrimination task, then the same manipulations produced large deficits suggesting that switching from one task to the other …
Reviewer #1 (Public Review):
The role of the parietal (PPC), the retrospenial (RSP) and the the visual cortex (S1) was assessed in three tasks corresponding a simple visual discrimination task, a working-memory task and a two-armed bandit task all based on the same sensory-motor requirements within a virtual reality framework. A differential involvement of these areas was reported in these tasks based on the effect of optogenetic manipulations. Photoinhibition of PPC and RSP was more detrimental than photoinhibition of S1 and more drastic effects were observed in presumably more complex tasks (i.e. working-memory and bandit task). If mice were trained with these more complex tasks prior to training in the simple discrimination task, then the same manipulations produced large deficits suggesting that switching from one task to the other was more challenging, resulting in the involvement of possibly larger neural circuits, especially at the cortical level. Calcium imaging also supported this view with differential signaling in these cortical areas depending on the task considered and the order to which they were presented to the animals. Overall the study is interesting and the fact that all tasks were assessed relying on the same sensory-motor requirements is a plus, but the theoretical foundations of the study seems a bit loose, opening the way to alternate ways of interpreting the data than "training history".
- Theoretical framework:
The three tasks used by the authors should be better described at the theoretical level. While the simple task can indeed be considered a visual discrimination task, the other two tasks operationally correspond to a working-memory task (i.e. delay condition which is indeed typically assessed in a Y- or a T-maze in rodent) or a two-armed bandit task (i.e. the switching task), respectively. So these three tasks are qualitatively different, are therefore reliant on at least partially dissociable neural circuits and this should be clearly analyzed to explain the rationale of the focus on the three cortical regions of interest. For the working-memory task we do not know the duration of the delay but this really is critical information; per definition, performance in such a task is delay-dependent, this is not explored in the paper.
Also, the authors heavily rely on "decision-making" but I am genuinely wondering if this is at all needed to account for the behavior exhibited by mice in these tasks (it would be more accurate for the bandit task) as with the perspective developed by the authors, any task implies a "decision-making" component, so that alone is not very informative on the nature of the cognitive operations that mice must compute to solve the tasks. I think a more accurate terminology in line with the specific task considered should be employed to clarify this.
The "switching"/bandit task is particularly interesting. But because the authors only consider trials with highest accuracy, I think they are missing a critical component of this task which is the balance between exploiting current knowledge and the necessity to explore alternate options when the former strategy is no longer effective. So trials with poor performance are thus providing an essential feedback which is a major drive to support exploratory actions and a critical asset of the bandit task. There is an ample literature documenting how these tasks assess the exploration/exploitation trade-off.
- Training history vs learning sets vs behavioral flexibility:
The authors consider "training history" as the unique angle to interpret the data. Because the experimental setup is the same throughout all experiments, I am wondering if animals are just simply provided with a cognitive challenge assessing behavioral flexibility given that they must identify the new rule while restraining from responding using previously established strategies. According to this view, it may be expected for cortical lesions to be more detrimental because multiple cognitive processes are now at play.
It is also possible that animals form learning sets during successive learning episodes which may interfere with or facilitate subsequent learning. Little information is provided regarding learning dynamics in each task (e.g. trials to criterion depending on the number of tasks already presented) to have a clear view on that.
- Calcium imaging data versus interventions:
The value of the calcium imaging data is not entirely clear. Does this approach bring a new point to consider to interpret or conclude on behavioral data or is it to be considered convergent with the optogenetic interventions? Very specific portions of behavioral data are considered for these analyses (e.g. only highly successful trials for the switching/bandit task) and one may wonder if considering larger or different samples would bring similar insights. The whole take on noise correlation is difficult to apprehend because of the same possible interpretation issue, does this really reflect training history, or that a new rule now must be implemented or something else? I don't really get how this correlative approach can help to address this issue.
- Theoretical framework:
-
Reviewer #2 (Public Review):
The authors use a combination of optogenetics and calcium imaging to assess the contribution of cortical areas (posterior parietal cortex, retrosplenial cortex, S1/V1) on a visual-place discrimination task. Head-fixed mice were trained on a simple version of the task where they were required to turn left or right depending on the visual cue that was present (e.g. X = go left; Y = go right). In a more complex version of the task the configurations were either switched during training or the stimuli were only presented at the beginning of the trial (delay).
The authors found that inhibiting the posterior parietal cortex and retrosplenial cortex affected performance, particularly on the complex tasks. However, previous training on the complex tasks resulted in more pronounced impairments on the simple task than …
Reviewer #2 (Public Review):
The authors use a combination of optogenetics and calcium imaging to assess the contribution of cortical areas (posterior parietal cortex, retrosplenial cortex, S1/V1) on a visual-place discrimination task. Head-fixed mice were trained on a simple version of the task where they were required to turn left or right depending on the visual cue that was present (e.g. X = go left; Y = go right). In a more complex version of the task the configurations were either switched during training or the stimuli were only presented at the beginning of the trial (delay).
The authors found that inhibiting the posterior parietal cortex and retrosplenial cortex affected performance, particularly on the complex tasks. However, previous training on the complex tasks resulted in more pronounced impairments on the simple task than when behaviourally naïve animals were trained/tested on a simple task. This suggests that the more complex tasks recruit these cortical areas to a greater degree, potentially due to increased attention required during the tasks. When animals then perform the simple version of the task their previous experience of the complex tasks is transferred to the simple task resulting in a different pattern of impairments compared to that found in behaviorally naïve animals.
The calcium imaging data showed a similar pattern of findings to the optogenetic study. There was overall increased activity in the switching tasks compared to the simple tasks consistent with the greater task demands. There was also greater trial-type selectivity in the switching task compared to the simple task. This increased trial-type selectivity in the switching tasks was subsequently carried forward to the simple task so that activity patterns were different when animals performed the simple task after experiencing the complex task compared to when they were trained on the simple task alone
Strengths:
The use of optogenetics and calcium-imaging enables the authors to look at the requirement of these brain structures both in terms of necessity for the task when disrupted as well as their contribution when intact.
The use of the same experimental set up and stimuli can provide a nice comparison across tasks and trials.
The study nicely shows that the contribution of cortical regions varies with task demands and that longer-term changes in neuronal responses c can transfer across tasks.
The study highlights the importance of considering previous experience and exposure when understanding behavioural data and the contribution of different regions.
The authors include a number of important controls that help with the interpretation of the findings.
Weaknesses:
There are some experimental details that need to be clarified to help with understanding the paper in terms of behavior and the areas under investigation.
The use of the same stimuli throughout is beneficial as it allows direct comparisons with animals experiencing the same visual cues. However, it does limit the extent to which you can extrapolate the findings. It is perhaps unsurprising to find that learning about specific visual cues affects subsequent learning and use of those specific cues. What would be interesting to know is how much of what is being shown is cue specific learning or whether it reflects something more general, for example schema learning which could be generalised to other learning situations. If animals were then trained on a different discrimination with different stimuli would this previous training modify behavior and neural activity in that instance. This would perhaps be more reflective of the types of typical laboratory experiments where you may find an impairment on a more complex task and then go on to rule out more simple discrimination impairments. However, this would typically be done with slightly different stimuli so you don't introduce transfer effects.
It is not clear whether length of training has been taken into account for the calcium imaging study given the slow development of neural representations when animals acquire spatial tasks.
The authors are presenting the study in terms of decision-making, however, it is unclear from the data as presented whether the findings specifically relate to decision making. I'm not sure the authors are demonstrating differential effects at specific decision points.
-
Reviewer #3 (Public Review):
Previous research has sought to understand the correlation between neuronal activity and decision-making in different regions of the cortex, and a plethora of cortical regions have been tested for necessity in a variety of decision-making tasks. For example, the posterior parietal cortex (PPC) has been shown to be necessary for visual decision-making tasks, whereas the retrosplenial cortex (RSC) is important in navigational planning. Although the necessity of different cortical regions in completion of certain discrimination tasks has yielded insights into these brain regions' roles, the previous experiences of each individual animal has not been explored. This raises the possibility that the previous experience and learned associations may affect how cortical areas process subsequent decision-making tasks.
T…
Reviewer #3 (Public Review):
Previous research has sought to understand the correlation between neuronal activity and decision-making in different regions of the cortex, and a plethora of cortical regions have been tested for necessity in a variety of decision-making tasks. For example, the posterior parietal cortex (PPC) has been shown to be necessary for visual decision-making tasks, whereas the retrosplenial cortex (RSC) is important in navigational planning. Although the necessity of different cortical regions in completion of certain discrimination tasks has yielded insights into these brain regions' roles, the previous experiences of each individual animal has not been explored. This raises the possibility that the previous experience and learned associations may affect how cortical areas process subsequent decision-making tasks.
To test this hypothesis, the authors used in vivo optogenetic activation of GABAergic interneurons to silence excitatory activity in the PPC, RSC, and S1 (a control, as S1 has not been shown to be involved in visual decision-making). They also employed 2-photon in vivo calcium imaging in head-fixed mice. A virtual Y-maze enabled the mice to decide to turn left or right to receive a reward. In the "simple task," mice had one rule - horizontal or vertical bars indicating whether the mouse should turn left or right to receive the reward. In the "complex task," there were two rules, A and B. Rule A was the exact same parameters as the simple task, whereas Rule B switched the left/right reward association with the horizontal/vertical bars. In some cases, there was an additional "complex task" where there was a delay between cue onset and decision making.
Inhibition of the PPC or RSC during the simple task resulted in small decreases in performance. Interestingly, inhibition of the PPC or RSC during more complex tasks, such as the delay task or switching task, resulted in much greater decrease in correct decisions. S1 inhibition decreased performance in the complex tasks, but not the simple task. Interestingly, when the animal was trained on a complex task (either the delay task or switching task) prior to testing in the simple task, there was a greater decrease in performance upon inhibition of the PPC or RSC compared to mice that had only undergone the simple task, implying that the prior experience of the complex task altered the cortical requirements for performing the simple task.
The next question was whether the neural activity was different in these cortical regions between tasks and between mice with and without previous experience. Neural activity in both the PPC and RSC was greater in mice that had solely undergone the switching task compared to mice that had only undergone the simple task. The trial-type selectivity of neurons was also higher in the switching task in both the PPC and RSC, and it took fewer cells to decode a trial accurately in mice from the switching task. Interestingly, compared to mice who had only experienced the simple task, animals that had prior experience with the switching task showed greater neuronal activity and neuronal selectivity in the PPC and RSC while performing the simple task.
-

-
