Environmental fluctuations reshape an unexpected diversity-disturbance relationship in a microbial community
Curation statements for this article:-
Curated by eLife

Evaluation Summary:
How species diversity responds to external perturbations (such as resource influxes and dilutions) is an important ecological question. Using soil microbial communities in devices where perturbations can be introduced in various forms and mathematical modelling, this study nicely illustrates how diversity is influenced by perturbations.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Environmental disturbances have long been theorized to play a significant role in shaping the diversity and composition of ecosystems. However, an inability to specify the characteristics of a disturbance experimentally has produced an inconsistent picture of diversity-disturbance relationships (DDRs). Here, using a high-throughput programmable culture system, we subjected a soil-derived bacterial community to dilution disturbance profiles with different intensities (mean dilution rates), applied either constantly or with fluctuations of different frequencies. We observed an unexpected U-shaped relationship between community diversity and disturbance intensity in the absence of fluctuations. Adding fluctuations increased community diversity and erased the U-shape. All our results are well-captured by a Monod consumer resource model, which also explains how U-shaped DDRs emerge via a novel ‘niche flip’ mechanism. Broadly, our combined experimental and modeling framework demonstrates how distinct features of an environmental disturbance can interact in complex ways to govern ecosystem assembly and offers strategies for reshaping the composition of microbiomes.
Article activity feed
-

-

Author Response:
Reviewer #2 (Public Review):
This work uses a throughput continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
A major strength of the work is a blending of modeling and empirical approaches. It includes an ambitiously-designed study that uses a controlled, high-throughput …
Author Response:
Reviewer #2 (Public Review):
This work uses a throughput continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
A major strength of the work is a blending of modeling and empirical approaches. It includes an ambitiously-designed study that uses a controlled, high-throughput microbial community experimental system to observe disturbance outcomes and uses those observations to build their proposed quantitative framework. The figures are informative and framework is explained clearly. The authors propose and name a new mechanism, "niche-flip" that describes resource competition at varying disturbance "intensities" - this is an interesting proposal and I suggest that it is explored more fully as a potential mechanism (see weaknesses).
Weaknesses of the work are the use of definitions that are generally inconsistent with the disturbance ecology literature, and the inability to separate the disturbance event characteristic of "intensity" from the biological outcome of mortality. The authors conclude that DDRs are contextual, which is supported by their modeling and data, but I suggest that they consider that diversity as an outcome in itself may not be the most informative metric of what mechanism(s) drive context-specific outcomes. The authors have a lot of compositional data that could also be examined to understand whether their "niche-flip" mechanism is supported.
This work is likely to advance our understanding of the myriad of outcomes of DDR and what potential mechanisms may support those DDR in natural ecosystems.
Thank you for your kind words and careful review of our manuscript. We are pleased you appreciate both the experiments and the modeling work, and that you are intrigued by the findings and the niche flip mechanism.
Major comments:
Comment 1. Ecological definitions and interdependence of disturbance outcomes/attributes
The authors define disturbance "intensity" as the average mortality rate but claim that this is a disturbance characteristic. However, mortality rate is not a characteristic of a disturbance event, but rather an effect/outcome of a disturbance on the biological community. The key distinction is that disturbance characteristics (also called traits or aspects) are defined relative to the environment, while disturbance outcomes (also called effects, impacts, or responses) are defined relative to the biology of interest, in this case a microbial community. So, changes in diversity of the community, as a result of a disturbance, is a biological outcome of the disturbance. An average mortality rate, what the authors call "intensity" (L40) would be such an outcome.
Thank you for this excellent point. We have revised the introduction to make this distinction, reproduced here for convenience:
"Accordingly, there have been many efforts aimed at understanding the role of environmental disturbances, which are perturbations to the state of an environment. These disturbances are of ecological interest for the impact they have on a community, for example, by bringing about mortality of organisms and a reduction of biomass of a community."
The authors' definition of "intensity" is not in agreement with the disturbance ecology literature, including the references cited in this current work. For example, in reference #18 (Miller et al. 2011 PNAS) disturbance aspects include intensity, timing, duration, extent, and interval. Specifically, Miller et al. 2011 defined intensity as the magnitude of the disturbance (e.g., a flood's maximum stage). Notably, Miller's definition of intensity is more aligned with the author's definition of "fluctuation," which the authors define as the "magnitude of deviations from the average". In the current work, the disturbance "event" cannot be separated from the biological outcome because of the nature of the continuous culture system. The system is not being disturbed with, for example, a change in pH or salinity or another environmental variable that results in microbial mortality, but rather the loss of viable members from the community through control of the flow-through. So, the mortality is both the precisely controlled disturbance "event" and "outcome" in the continuous culture.
To summarize, the premise of the article is confusing, because one of the two disturbance "characteristics" considered is, rather a disturbance outcome. This may seem like mincing words and to each paper its own definitions, but because this work seeks to reconcile DDRs as reported across many studies, and because many of the previous ecology studies that have investigated or reported DDRs are not using analogous terms, the work could further confusion rather than serve as a reconciliation. When different definitions are applied that mix disturbance aspects with biological outcomes of disturbance, readers will have to work hard to understand this work in context with the existing literature. I suggest revising the introductory section to be consistent in terminology with the ecology literature and to be framed not only as disturbance characteristics, but also outcomes. I also suggest adding discussion of how an inability to distinguish disturbance event from outcome may influence interpretation of this work and its broader application. I suggest adding clarification/discussion of "how intensity and fluctuations interact" (e.g. L200): as the authors define intensity and fluctuation of the disturbance event, intensity is not independent of the biological disturbance outcome of mortality in the given model system. So, how the two "disturbance components interact" is not able to be examined independently from the biological outcome (mortality, resulting diversity).
These are also critical points. First, we will address the choice of terminology (re: Miller et al) and, second, the equivalence between disturbance and outcome in continuous culture.
We agree that careful use of terminology is important for understanding our work in context of the literature. Accordingly, we have replaced our characteristics “intensity” and “fluctuation” with “mean intensity” and “frequency” throughout the paper. We have also added more examples through the results section to indicate how mean intensity, frequency, and maximum dilution rates (during disturbance events) are related.
"To determine whether the effects of disturbance on diversity are truly fluctuation-dependent15, a disturbance should ideally be decomposed into distinct components of mean intensity (e.g. time-averaged disturbance magnitude) and frequency (e.g. temporal profile of fluctuations)."
The direct connection between disturbance and mortality in a continuous culture system under dilution disturbances is a critical aspect of our experimental design, because we wanted to compare disturbance outcomes that varied in temporal features (in Miller et al terms, intensity/magnitude vs frequency/timing) while holding mortality equal. In continuous culture this may be achieved by controlling dilution rate and frequency, but you are correct that other classes of natural disturbances such as pH or salinity changes may have different effects on community members. As a first step towards investigating these effects, we had included analyses with non-equal mortality rates (Appendix figure 4). We have now edited the introduction and discussion to emphasize that the equivalence between disturbance event and disturbance outcome is a feature specific to continuous culture.
Introduction
"Dilution is perhaps the most common choice for a laboratory disturbance, as it causes species-independent mortality and replenishes the system with fresh nutrients, reminiscent of flow in soil, aquatic, or gut microbiomes. Unlike disturbances with indirect biological impacts (such as pH, temperature, or osmolarity disturbances), there is a direct link between the dilution disturbance event (removal of culture volume) and the biological outcome (mortality of community members)."
Discussion
"We also note however, that these types of disturbances do not share the direct link between environmental change and biological outcome that is characteristic of dilution disturbance, so the impact may be less clear."
Comment 2: Compositional evidence for the proposed "niche flip" mechanism and suggestion for deeper consideration of population-level response to disturbance outcomes that collectively contribute to emergent diversity values.
Regarding the "niche flip" - it is unclear whether there is compositional evidence for any swap in niche preference/space among particular community members. Figure S8 may offer evidence, but I could not deduce it from the busy bar charts. Could population/ASV level analysis be conducted on each member to assess their dynamics and ask whether the dynamics support the proposed niche-flip as a DDR mechanism?
This is a very interesting suggestion. As suggested, we could extract the relative preferences of different ASVs from composition data to test a prediction about changes in the composition resulting from niche flip. To make such a prediction, we’d need the Monod growth parameters of the species on relevant resources. We began collecting this data (see Figure 3 – figure supplement 4) but found it challenging to measure these parameters on defined media sources. Furthermore, since we elected to run our main experiments in a complex media that could potentially support diverse communities (as opposed to minimal medias which produce simple communities, see Goldford et al Science 2018) we cannot link Monod growth parameters in this media to particular resources. Subsequent experiments with defined species with measured Monod parameters in defined media would enable us to make and test predictions. These are sizeable experiments that we do not believe are in the scope of the present work. Without a testable prediction, we do not believe species or ASV level analysis to be particularly informative on its own.
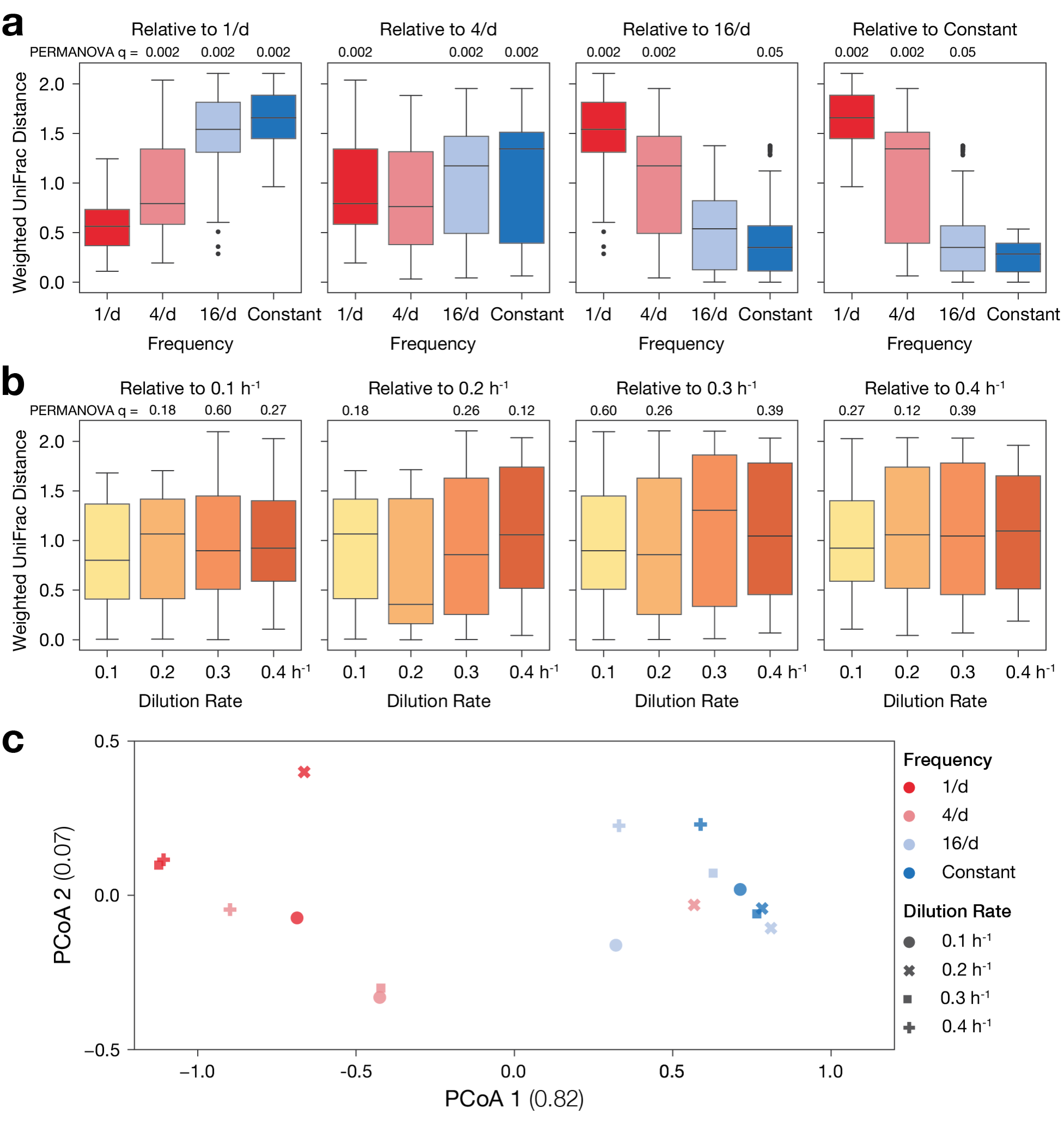
Related, there seems to be possible evidence of a "fluctuation" rate threshold, after which there is a major compositional shift in the microbial community. Consider Figure 3: At all "intensities", there is a shift in microbial community composition between "fluctuation" rates of 4/day and 16/day (3d, Fig S8). This threshold/shift is not also apparent in the Shannon diversity in Fig 3f. This could be an example in which diversity as a metric in itself is not as informative/useful outcome for disturbance responses, as identical Shannon diversity values can result from different community compositions that are themselves the outcomes of different mechanisms. I see from the PCoAs (Fig S9) that the authors were exploring potential compositional clustering by day, frequency, and dilution - the most "obvious" clustering to the eye is indeed by "frequency" and between 4/day and 16/day (red/blue separation along both axes, which also supports a potential threshold/shift. Generally, it would have been good to report statistical tests (e.g., PERMANOVA or equivalent) for these PCoA categories (where it makes sense, nested and term interactions as well) - is there statistical support for compositional threshold shift between 4/16?
Thank you for these suggestions. Indeed, by eye and by the PCoA plots, there seems to be a significant difference in composition that separate the low-frequency (1/day & 4/day) from the high-frequency (16/day & Constant) conditions. We calculated pairwise distances between Day 6 samples grouped by A) dilution frequency, B) mean dilution rate, or C) combinations of dilution rate and frequency. Using these distances to perform PERMANOVA tests, we find significant differences between cultures with different frequencies, but not for cultures with different dilution rates. For combinations, we found several pairs with differences that were significant only before correction for false-discovery rate. Distances between low-frequency (1/day & 4/day) conditions are much smaller than between low-frequency and high-frequency groups, or between the high-frequency groups. We have now included this as Figure 3 – figure supplement 9 and have summarized the results in the main text, reproduced below for convenience:
"PERMANOVA statistical analysis of endpoint compositions confirmed that dilution frequency (but not mean dilution rate) had a significant effect on composition (Figure 3 – figure supplement 9). Despite separation between conditions in PCoA of endpoint compositions (Figure 3 – figure supplement 9), PERMANOVA analysis of dilution rate and frequency combinations did not yield significant values after correcting for false discovery rate."
Reviewer #3 (Public Review):
This manuscript focuses on the relationship between diversity and disturbance. The authors study this relationship in experimental microbial communities. These communities as subject to different levels of disturbance, which is identified as the dilution rate. The authors find a non-monotonic relationship between diversity and dilution rate. In presence of temporal fluctuations, the non-monotonic relationship becomes less evident, disappearing for strong enough fluctuations. The experimental findings are well explained by a consumer-resource model with Monod response.
The results of the paper are a very interesting combination of experimental and theoretical work. The manuscript is well written and easy to follow.
Experiments. The data support the main result of the paper. The U-shaped disturbance-diversity relationship (DDR) is robust (e.g., independent of the measure of diversity). The experimental setup is innovative.
Theory. A main strength of the manuscript is the clarity in which the model reproduces the experimental data. It is also interesting that alternative models (Lotka-Volterra and consumer-resource with linear response) do not reproduce the data, therefore indicating the relevance of the data themselves. The main weakness of the paper is that, in the end, the mechanism behind the non-monotonicity of the DDR is not completely clear. The authors discuss how it emerges with two species and two resources in presence of a trade-off between maximal growth rate and resource-limited growth rate: at low dilution rate, the species with high maximal growth rate wins, while at high dilution rate the one with resource-limited growth rate dominates. This mechanism is clear with two species (in which diversity can transition between 2 and 1). It is unclear what happens for more species and resources. In particular, the role of the tradeoff --- which is central in the pairwise competition case --- is unclear: the U-shapes relationship is observed also in absence of the tradeoff for multispecies communities.
Thank you for your enthusiasm about our work and your careful review of our manuscript. We are pleased you appreciate the concordance between experiment and model in our study.
-
Evaluation Summary:
How species diversity responds to external perturbations (such as resource influxes and dilutions) is an important ecological question. Using soil microbial communities in devices where perturbations can be introduced in various forms and mathematical modelling, this study nicely illustrates how diversity is influenced by perturbations.
(This preprint has been reviewed by eLife. We include the public reviews from the reviewers here; the authors also receive private feedback with suggested changes to the manuscript. Reviewer #1 agreed to share their name with the authors.)
-
Reviewer #1 (Public Review):
This work uses a continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
Authors cultured a soil microbial community in chemostats with varying dilution rates (disturbances) and surveyed species abundances at the steady state. At a constant dilution rate, an U shaped DDR …
Reviewer #1 (Public Review):
This work uses a continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
Authors cultured a soil microbial community in chemostats with varying dilution rates (disturbances) and surveyed species abundances at the steady state. At a constant dilution rate, an U shaped DDR (diversity-disturbance relationship) is observed: diversity is the lowest at intermediate dilution rate. When dilution rate is allowed to fluctuate, increased diversity is observed. Moreover, the U-chape is erased when fluctuation in dilution rate is very large (one large dilution per day instead of many smaller dilutions per day). These phenomena can be explained by a resource competition mathematical model where growth rate is a saturable function of resource, but not by a Lotka-Volterra model or if growth rate is a linear function of resource. Finally, by simulating perturbations at different fluctuation frequencies, authors obtained diverse DDRs including U shape, peaked, increasing, decreasing, and flat.
A major strength of the work is a blending of modeling and empirical approaches. The figures are informative and framework is explained clearly.
-
Reviewer #2 (Public Review):
This work uses a throughput continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
A major strength of the work is a blending of modeling and empirical approaches. It includes an ambitiously-designed study that uses a controlled, high-throughput microbial community …
Reviewer #2 (Public Review):
This work uses a throughput continuous culture system with simplified soil microbial communities to investigate how diversity-disturbance relationships (DDRs) change with different disturbance "intensities" (here, defined as mortality rate or dilution rate in a continuous system) and "frequencies" (here, defined as the number of dilution events that occur per day to achieve the desired mortality rate). Understanding the mechanisms that support different DDR is an ongoing and urgent need in ecology and ecosystem sciences because of the pressing need to predict and manage systems given climate and land-use disturbances.
A major strength of the work is a blending of modeling and empirical approaches. It includes an ambitiously-designed study that uses a controlled, high-throughput microbial community experimental system to observe disturbance outcomes and uses those observations to build their proposed quantitative framework. The figures are informative and framework is explained clearly. The authors propose and name a new mechanism, "niche-flip" that describes resource competition at varying disturbance "intensities" - this is an interesting proposal and I suggest that it is explored more fully as a potential mechanism (see weaknesses).
Weaknesses of the work are the use of definitions that are generally inconsistent with the disturbance ecology literature, and the inability to separate the disturbance event characteristic of "intensity" from the biological outcome of mortality. The authors conclude that DDRs are contextual, which is supported by their modeling and data, but I suggest that they consider that diversity as an outcome in itself may not be the most informative metric of what mechanism(s) drive context-specific outcomes. The authors have a lot of compositional data that could also be examined to understand whether their "niche-flip" mechanism is supported.
This work is likely to advance our understanding of the myriad of outcomes of DDR and what potential mechanisms may support those DDR in natural ecosystems.
Major comments:
Comment 1. Ecological definitions and interdependence of disturbance outcomes/attributes
The authors define disturbance "intensity" as the average mortality rate but claim that this is a disturbance characteristic. However, mortality rate is not a characteristic of a disturbance event, but rather an effect/outcome of a disturbance on the biological community. The key distinction is that disturbance characteristics (also called traits or aspects) are defined relative to the environment, while disturbance outcomes (also called effects, impacts, or responses) are defined relative to the biology of interest, in this case a microbial community. So, changes in diversity of the community, as a result of a disturbance, is a biological outcome of the disturbance. An average mortality rate, what the authors call "intensity" (L40) would be such an outcome.
The authors' definition of "intensity" is not in agreement with the disturbance ecology literature, including the references cited in this current work. For example, in reference #18 (Miller et al. 2011 PNAS) disturbance aspects include intensity, timing, duration, extent, and interval. Specifically, Miller et al. 2011 defined intensity as the magnitude of the disturbance (e.g., a flood's maximum stage). Notably, Miller's definition of intensity is more aligned with the author's definition of "fluctuation," which the authors define as the "magnitude of deviations from the average". In the current work, the disturbance "event" cannot be separated from the biological outcome because of the nature of the continuous culture system. The system is not being disturbed with, for example, a change in pH or salinity or another environmental variable that results in microbial mortality, but rather the loss of viable members from the community through control of the flow-through. So, the mortality is both the precisely controlled disturbance "event" and "outcome" in the continuous culture.
To summarize, the premise of the article is confusing, because one of the two disturbance "characteristics" considered is, rather a disturbance outcome. This may seem like mincing words and to each paper its own definitions, but because this work seeks to reconcile DDRs as reported across many studies, and because many of the previous ecology studies that have investigated or reported DDRs are not using analogous terms, the work could further confusion rather than serve as a reconciliation. When different definitions are applied that mix disturbance aspects with biological outcomes of disturbance, readers will have to work hard to understand this work in context with the existing literature. I suggest revising the introductory section to be consistent in terminology with the ecology literature and to be framed not only as disturbance characteristics, but also outcomes. I also suggest adding discussion of how an inability to distinguish disturbance event from outcome may influence interpretation of this work and its broader application. I suggest adding clarification/discussion of "how intensity and fluctuations interact" (e.g. L200): as the authors define intensity and fluctuation of the disturbance event, intensity is not independent of the biological disturbance outcome of mortality in the given model system. So, how the two "disturbance components interact" is not able to be examined independently from the biological outcome (mortality, resulting diversity).
Comment 2: Compositional evidence for the proposed "niche flip" mechanism and suggestion for deeper consideration of population-level response to disturbance outcomes that collectively contribute to emergent diversity values.
Regarding the "niche flip" - it is unclear whether there is compositional evidence for any swap in niche preference/space among particular community members. Figure S8 may offer evidence, but I could not deduce it from the busy bar charts. Could population/ASV level analysis be conducted on each member to assess their dynamics and ask whether the dynamics support the proposed niche-flip as a DDR mechanism?
Related, there seems to be possible evidence of a "fluctuation" rate threshold, after which there is a major compositional shift in the microbial community. Consider Figure 3: At all "intensities", there is a shift in microbial community composition between "fluctuation" rates of 4/day and 16/day (3d, Fig S8). This threshold/shift is not also apparent in the Shannon diversity in Fig 3f. This could be an example in which diversity as a metric in itself is not as informative/useful outcome for disturbance responses, as identical Shannon diversity values can result from different community compositions that are themselves the outcomes of different mechanisms. I see from the PCoAs (Fig S9) that the authors were exploring potential compositional clustering by day, frequency, and dilution - the most "obvious" clustering to the eye is indeed by "frequency" and between 4/day and 16/day (red/blue separation along both axes, which also supports a potential threshold/shift. Generally, it would have been good to report statistical tests (e.g., PERMANOVA or equivalent) for these PCoA categories (where it makes sense, nested and term interactions as well) - is there statistical support for compositional threshold shift between 4/16?
-
Reviewer #3 (Public Review):
This manuscript focuses on the relationship between diversity and disturbance. The authors study this relationship in experimental microbial communities. These communities as subject to different levels of disturbance, which is identified as the dilution rate. The authors find a non-monotonic relationship between diversity and dilution rate. In presence of temporal fluctuations, the non-monotonic relationship becomes less evident, disappearing for strong enough fluctuations. The experimental findings are well explained by a consumer-resource model with Monod response.
The results of the paper are a very interesting combination of experimental and theoretical work. The manuscript is well written and easy to follow.
Experiments. The data support the main result of the paper. The U-shaped disturbance-diversity …
Reviewer #3 (Public Review):
This manuscript focuses on the relationship between diversity and disturbance. The authors study this relationship in experimental microbial communities. These communities as subject to different levels of disturbance, which is identified as the dilution rate. The authors find a non-monotonic relationship between diversity and dilution rate. In presence of temporal fluctuations, the non-monotonic relationship becomes less evident, disappearing for strong enough fluctuations. The experimental findings are well explained by a consumer-resource model with Monod response.
The results of the paper are a very interesting combination of experimental and theoretical work. The manuscript is well written and easy to follow.
Experiments. The data support the main result of the paper. The U-shaped disturbance-diversity relationship (DDR) is robust (e.g., independent of the measure of diversity). The experimental setup is innovative.
Theory. A main strength of the manuscript is the clarity in which the model reproduces the experimental data. It is also interesting that alternative models (Lotka-Volterra and consumer-resource with linear response) do not reproduce the data, therefore indicating the relevance of the data themselves. The main weakness of the paper is that, in the end, the mechanism behind the non-monotonicity of the DDR is not completely clear. The authors discuss how it emerges with two species and two resources in presence of a trade-off between maximal growth rate and resource-limited growth rate: at low dilution rate, the species with high maximal growth rate wins, while at high dilution rate the one with resource-limited growth rate dominates. This mechanism is clear with two species (in which diversity can transition between 2 and 1). It is unclear what happens for more species and resources. In particular, the role of the tradeoff --- which is central in the pairwise competition case --- is unclear: the U-shapes relationship is observed also in absence of the tradeoff for multispecies communities.
-
