Development of frequency tuning shaped by spatial cue reliability in the barn owl’s auditory midbrain
Curation statements for this article:-
Curated by eLife

eLife assessment
This is a valuable study that uses an innovative approach to show how the statistics of sound stimulation influence neural coding in an experience-dependent fashion in the developing barn owl brain. The paper will therefore be of interest to the fields of developmental and sensory neuroscience. In its present form, however, the evidence presented to support the main claims of the paper is incomplete and could be strengthened in various ways.
This article has been Reviewed by the following groups
Discuss this preprint
Start a discussion What are Sciety discussions?Listed in
- Evaluated articles (eLife)
Abstract
Sensory systems preferentially strengthen responses to stimuli based on their reliability at conveying accurate information. While previous reports demonstrate that the brain reweighs cues based on dynamic changes in reliability, how the brain may learn and maintain neural responses to sensory statistics expected to be stable over time is unknown. The barn owl’s midbrain features a map of auditory space where neurons compute horizontal sound location from the interaural time difference (ITD). Frequency tuning of midbrain map neurons correlates with the most reliable frequencies for the neurons’ preferred ITD (Cazettes et al., 2014). Removal of the facial ruff led to a specific decrease in the reliability of high frequencies from frontal space. To directly test whether permanent changes in ITD reliability drive frequency tuning, midbrain map neurons were recorded from adult owls, with the facial ruff removed during development, and juvenile owls, before facial ruff development. In both groups, frontally tuned neurons were tuned to frequencies lower than in normal adult owls, consistent with the change in ITD reliability. In addition, juvenile owls exhibited more heterogeneous frequency tuning, suggesting normal developmental processes refine tuning to match ITD reliability. These results indicate causality of long-term statistics of spatial cues in the development of midbrain frequency tuning properties, implementing probabilistic coding for sound localization.
Article activity feed
-

-

Author Response
Reviewer #1 (Public Review):
- It would be helpful to include some sort of comparison in Fig. 4, e.g. the regressions shown in Fig 3, to indicate to what extent the ICCl data corresponds to the "control range" of frequency tuning.
Figure 4 was modified to show the frequency range typically found in the ICCls. This range is based on results from Wagner et al., 2007, which extensively surveyed ICCls responses. This modification shows that our ICCls recordings in the ruff-removed owls cover the normal frequency hearing range of the owl.
- A central hypothesis of the study is that the frequency preference of the high-frequency neurons is lower in ruff-removed owls because of the lowered reliability caused by a lack of the ruff. Yet, while lower, the frequency range of many neurons in juvenile and ruff-removed owls seems …
Author Response
Reviewer #1 (Public Review):
- It would be helpful to include some sort of comparison in Fig. 4, e.g. the regressions shown in Fig 3, to indicate to what extent the ICCl data corresponds to the "control range" of frequency tuning.
Figure 4 was modified to show the frequency range typically found in the ICCls. This range is based on results from Wagner et al., 2007, which extensively surveyed ICCls responses. This modification shows that our ICCls recordings in the ruff-removed owls cover the normal frequency hearing range of the owl.
- A central hypothesis of the study is that the frequency preference of the high-frequency neurons is lower in ruff-removed owls because of the lowered reliability caused by a lack of the ruff. Yet, while lower, the frequency range of many neurons in juvenile and ruff-removed owls seems sufficiently high to be still responsive at 7-8 kHz. I think it would be important to know to what extent neurons are still ITD sensitive at the "unreliable high frequencies" even if the CFs are lower since the "optimization" according to reliability depends not on the best frequency of each neuron per se, but whether neurons are less ITD sensitive at the higher, less reliable frequencies.
The concern regarding the frequency range that elicits responsivity was largely addressed above. Specifically, Figure L1 showing frequency tuning of frontally tuned ICx neurons in ruff-removed owls indicates that while there is some variability of tuning across neurons, there is little responsivity above 6 kHz. In contrast, equivalent analysis in juvenile owls (Figure L3), shows there is much more responsiveness and variability across neurons to high and low frequencies. This evidence supports our hypothesis that the juvenile owl brain is still highly plastic, which facilitates learning during development. Although the underlying data was already reported in Figure 7 of our previously submitted manuscript, we can include Figures L1 and L2, potentially as supplemental figures, if considered useful by editors and reviewers. Nevertheless, this argumentation was further expanded in the revised text (Line 229).
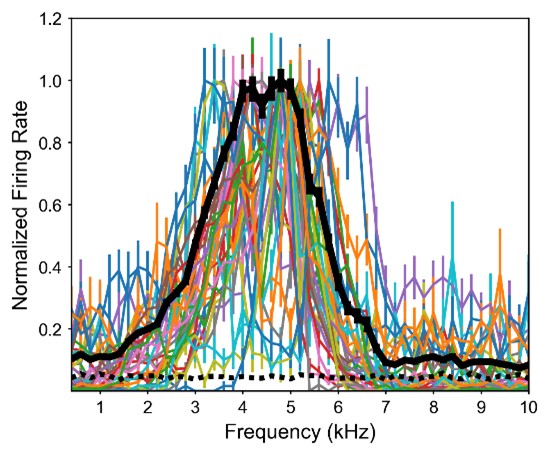
Figure L1. Frequency tuning of frontally-tuned ICx neurons in ruff-removed owls. Tuning curves are normalized by the max response. Thick black line indicates the average tuning curve. Dashed black line indicates basal response.
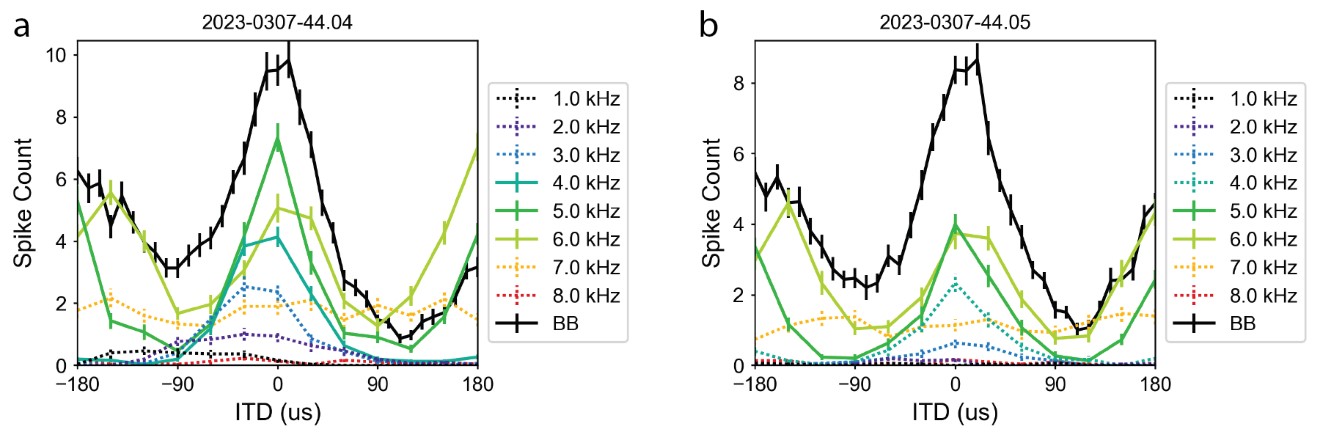
Figure L2. ITD sensitivity across frequencies in ruff-removed owl. Two example neurons shown in a and b. ITD tuning for tones (colored) and broadband (black) plotted by firing rate (non-normalized). Solid colored lines indicate responses to frequencies that are within the neuron’s preferred frequency range (i.e. above the half-height, see Methods), dashed lines indicate frequencies outside of the neuron’s frequency range.
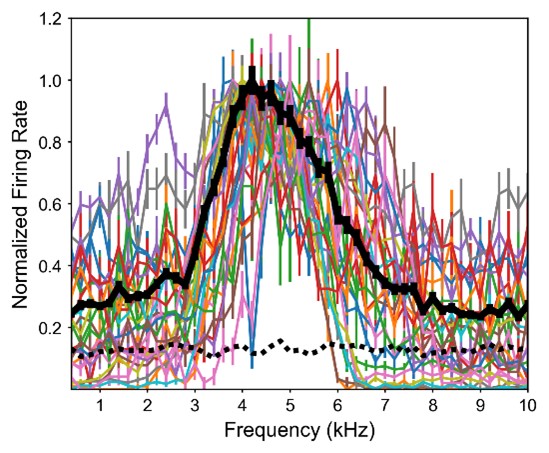
Figure L3. Frequency tuning of frontally-tuned ICx neurons in juvenile owls. Tuning curves are normalized by the max response. Thick black line indicates the average tuning curve. Dashed black line indicates basal response.
- It would be interesting to have an estimate of the time scale of experience dependency that induces tuning changes. Do the authors have any data on this question? I appreciate the authors' notion that the quantifications in Fig 7 might indicate that juvenile owls are already "beginning to be shaped by ITD reliability" (line 323 in Discussion). How many days after hearing onset would this correspond to? Does this mean that a few days will already induce changes?
While tracking changes induced by ruff-removal over development were outside of the scope of this study, many other studies have assessed experience-dependent plasticity in the barn owl. The recordings in this study were performed approximately 20 days after hearing onset, suggesting that the juveniles had ample time to begin learning. These points were expanded upon in the discussion (Lines 254, 280-283).
Reviewer #2 (Public Review):
- Why is IPD variability plotted instead of ITD variability (or indeed spatial reliability)? The relationship between these measures is likely to vary across frequency, which makes it difficult to compare ITD variability across frequency when IPDs are plotted. Normalizing data across frequencies also makes it difficult to compare different locations and acoustical conditions. For example, in Fig.1a and Fig.1b, the data shown for 3 kHz at ~160 degrees seems quantitatively and visually quite different, but the difference (in Fig.1c) appears to be negligible.
Justification of why IPD variability is used as an estimate of ITD variability was added to introduction (Lines 55-60), results (Line 100) and methods (Lines 371-374) sections of the manuscript, explaining the fact that because ITD detection is based on phase locking by auditory nerve and ITD detector neurons tuned to narrow frequency bands, responses of ITD detector neurons forwarded to downstream midbrain regions are therefore determined by IPD variability. Additionally, ITD is calculated by dividing IPD by frequency, which makes comparisons of ITD reliability across frequency mathematically uninformative.
- How well do the measures of ITD reliability used reflect real-world listening? For example, the model used to calculate ITD reliability appears to assume the same (flat) spectral profile for targets and distractors, which are presented simultaneously with the same temporal envelope, and a uniform spatial distribution of sounds across space. It is therefore unclear how robust the study's results are to violations of these assumptions.
While we agree that our analysis cannot completely capture real-world listening for the barn owl, a general analysis using similar flat spectral profiles for targets and concurrent sounds provides a broad assessment of reliability of ITD cues. While a full recapitulation of real-world listening is beyond the scope of this study (i.e. recording natural scenes from the ear canals of wild barn owls), we included additional analyses of ITD reliability in Figure 1-figure supplement 1, described above.
- Does facial ruff removal produce an isolated effect on ITD variability or does it also produce changes in directional gain, and the relationship between spatial cues and sound location? Although the study considers this issue in some places (e.g. Fig.2, Fig.5), a clearer presentation of the acoustical effects of facial ruff removal and their implications (for all locations, not just those to the front), as well as an attempt to understand how these acoustical changes lead to the observed changes in ITD reliability, would greatly strengthen the study. In addition, Fig.1 shows average ITD reliability across owls, but it would be helpful to know how consistent these measures are across owls, given individual variability in Head-Related Transfer Functions (HRTFs). This potentially has implications for the electrophysiological experiments, if the HRTFs of those animals were not measured. One specific question that is potentially very relevant is whether the facial ruff attenuates sounds presented behind the animal and whether it does so in a frequency-dependent way. In addition, if facial ruff removal enables ILDs to be used for azimuth, then ITDs may also become less necessary at higher frequencies, even if their reliability remains unchanged.
Additional analysis was conducted to generate representation of changes in directional gain induced by ruff removal, added to new figure (Fig 5). This analysis shows that changes in gain following ruff-removal are largely frequency-independent: there is a de-attenuation of peripherally and rearwardly located sounds, but the highest gain remains for high frequencies in frontal space. There is an additional increase in gain for high frequencies from rearward space, these changes would not explain the changes in frequency tuning we report. As mentioned in new additions to the manuscript, the changes at the most rearward-located auditory spatial locations are unlikely to have an effect on the auditory midbrain. No studies in the barn owl have found neurons in the ICx or optic tectum tuned to >120° (Knudsen, 1982; Knudsen, 1984; Cazettes et al., 2014). In addition, variability of IPD reliability across owls was analyzed and reported in the amended Figure 1, which notes very little changes across owls. In this analysis, we did realize that the file of one of the HRTFs obtained from von Campenhausen et al. 2006 was mislabeled, which explains slight differences in revised Fig 1b. Nevertheless, added analysis of IPD reliability across owls indicates that the pattern in ITD reliability is stable across owls (Fig. 1d,e), which supports our decision to not record HRTFs from owls used in this study. Finally, we added to the discussion that clarifies that the use of ILD for azimuth would not provide the same resolution as ITD would (Lines 295-303). We also do not believe that the use of ILD for azimuth would make “ITDs… less necessary at higher frequencies”, given that the ICCls is still computing ITD at these high frequencies (Fig 4), and that ILDs also have higher resolution at higher frequencies, with and without the facial ruff (Olsen et al, 1989; Keller et al., 1998; von Campenhausen et al., 2006).
- It is unclear why some analyses (Fig.5, Fig.7) are focused on frontal locations and frontally-tuned neurons. It is also unclear why neurons with a best ITDs of 0 are described as frontally tuned since locations behind the animal produce an ITD of 0 also. Related to this, in Fig.1, facial ruff removal appears to reduce IPD variability at low frequencies for locations to the rear (~160 degrees), where the ITD is likely to be close to 0. Neurons with a best ITD of 0 might therefore be expected to adjust their frequency tuning in opposite directions depending on whether they are tuned to frontal or rearward locations.
An extensive explanation was added to the methods detailing why we do not believe the neurons recorded in this study are tuned to the rear. Namely, studies mapping the barn owl’s ICx and optic tectum have not reported neurons tuned to locations >120°, with the number of neurons representing a given spatial location decreasing with eccentricity (Knudsen, 1982; Knudsen, 1984; Cazettes et al., 2014). While we agree that there does seem to be a change in ITD reliability at ~160° following ruff-removal, the result is largely similar to the change that occurs in frontal space (Fig 1b), which is consistent with the ruff-removed head functioning as a sphere. Thus, we wouldn’t expect rearwardly-tuned neurons, if they could be readily found, to adjust their frequency tuning to higher frequencies. Finally, we want to clarify that we focused our analyses on frontally-tuned neurons because frontal space is where we observed the largest change in ITD reliability. Text was added to the Discussion section to clarify this point (Lines 313-321).
- The study suggests that information about high-frequency ITDs is not passed on to the ICX if the ICX does not contain neurons that have a high best frequency. However, neurons might be sensitive to ITDs at frequencies other than the best frequency, particularly if their frequency tuning is broader. It is also unclear whether the best frequency of a neuron always corresponds to the frequency that provides the most reliable ITD information, which the study implicitly assumes.
The concern about ITD sensitivity at non-preferred frequencies was addressed under the essential revision #3, as well as under Reviewer 1’s concerns.
-
eLife assessment
This is a valuable study that uses an innovative approach to show how the statistics of sound stimulation influence neural coding in an experience-dependent fashion in the developing barn owl brain. The paper will therefore be of interest to the fields of developmental and sensory neuroscience. In its present form, however, the evidence presented to support the main claims of the paper is incomplete and could be strengthened in various ways.
-
Reviewer #1 (Public Review):
This study investigated how changes in spatial stimulus statistics affect neuronal tuning properties in the barn owl, a well-studied model organism of spatial processing and sound localization. The authors utilized the fact that the owls' facial ruff significantly affects the reliability of binaural cues at specific frequencies. To this end, they compared the tuning to frequency and interaural time differences (ITD) of midbrain (ICX) neurons in adult owls with intact or removed ruff and juvenile owls (with undeveloped ruff). They find that frequency preference is lowered at frontally tuned neurons in the absence of intact /fully developed facial ruff, in accordance with the notion that ITD reliability is lowered for higher frequencies by the lack of ruff. Likewise, they find that ITD tuning width is …
Reviewer #1 (Public Review):
This study investigated how changes in spatial stimulus statistics affect neuronal tuning properties in the barn owl, a well-studied model organism of spatial processing and sound localization. The authors utilized the fact that the owls' facial ruff significantly affects the reliability of binaural cues at specific frequencies. To this end, they compared the tuning to frequency and interaural time differences (ITD) of midbrain (ICX) neurons in adult owls with intact or removed ruff and juvenile owls (with undeveloped ruff). They find that frequency preference is lowered at frontally tuned neurons in the absence of intact /fully developed facial ruff, in accordance with the notion that ITD reliability is lowered for higher frequencies by the lack of ruff. Likewise, they find that ITD tuning width is increased in juvenile and ruff-removed owls, providing further indications for a lowered frequency preference (because ITD tuning width is correlated with wavelength). While the authors cannot provide causal evidence that ITD reliability is the driver for these experience-dependent changes, the data is very consistent with this interpretation. Thus, the conclusions are mostly well supported and will add interesting aspects to our understanding of spatial (ITD) coding and the role of stimulus statistics in general. Nonetheless, a few questions should be clarified that would strengthen the conclusions in my opinion:
- It would be helpful to include some sort of comparison in Fig. 4, e.g. the regressions shown in Fig 3, to indicate to what extent the ICCl data corresponds to the "control range" of frequency tuning.
- A central hypothesis of the study is that the frequency preference of the high-frequency neurons is lower in ruff-removed owls because of the lowered reliability caused by a lack of the ruff. Yet, while lower, the frequency range of many neurons in juvenile and ruff-removed owls seems sufficiently high to be still responsive at 7-8 kHz. I think it would be important to know to what extent neurons are still ITD sensitive at the "unreliable high frequencies" even if the CFs are lower since the "optimization" according to reliability depends not on the best frequency of each neuron per se, but whether neurons are less ITD sensitive at the higher, less reliable frequencies.
- It would be interesting to have an estimate of the time scale of experience dependency that induces tuning changes. Do the authors have any data on this question? I appreciate the authors' notion that the quantifications in Fig 7 might indicate that juvenile owls are already "beginning to be shaped by ITD reliability" (line 323 in Discussion). How many days after hearing onset would this correspond to? Does this mean that a few days will already induce changes?
-
Reviewer #2 (Public Review):
This study investigates whether frequency tuning in the avian auditory midbrain is changed by the reliability of a key sound localization cue (Interaural Time Differences, ITDs) during development. It tests whether auditory neurons become more sensitive to sound frequencies that provide more reliable information about ITDs.
To manipulate the reliability of ITDs in a frequency-specific way, the authors removed the facial ruff of barn owls during development, which alters the acoustical input available to the animal in a number of important ways. When these animals reached adulthood, electrophysiological recordings were performed in the external nucleus of the inferior colliculus (ICx). Compared to control animals, these recordings revealed a weaker relationship between the best-frequency and best-ITD of …
Reviewer #2 (Public Review):
This study investigates whether frequency tuning in the avian auditory midbrain is changed by the reliability of a key sound localization cue (Interaural Time Differences, ITDs) during development. It tests whether auditory neurons become more sensitive to sound frequencies that provide more reliable information about ITDs.
To manipulate the reliability of ITDs in a frequency-specific way, the authors removed the facial ruff of barn owls during development, which alters the acoustical input available to the animal in a number of important ways. When these animals reached adulthood, electrophysiological recordings were performed in the external nucleus of the inferior colliculus (ICx). Compared to control animals, these recordings revealed a weaker relationship between the best-frequency and best-ITD of individual neurons. A similarly weak relationship was observed in young animals whose ruff had not yet fully developed.
These results arise partly because animals without a facial ruff possess neurons with a best ITD of 0 that are tuned to unusually low frequencies. Having considered a number of possible explanations, the authors argue that this occurs because facial ruff removal reduces the reliability of high-frequency ITDs for frontal locations. Consequently, neurons tuned to frontal locations shift their frequency sensitivity to lower frequencies, which provides more reliable information about ITD. This shift toward lower frequencies is also thought to partly explain changes in tuning width that are observed in the absence of a facial ruff.
The study concludes that these results collectively provide evidence that the brain learns to implement probabilistic coding of sound location during development. However, although the study clearly shows changes in neural tuning in the absence of a fully developed facial ruff, the causal link with ITD reliability is complicated by a number of technical issues. The most important of these include a tendency to ignore the rear hemifield for some analyses but not others, the complex acoustical effects of facial ruff removal, and a model of IPD reliability that may or may not accurately reflect real-world listening. Nevertheless, the study presents an interesting set of results and shows an innovative approach in a number of places.
ACOUSTICS: A key strength of the study is its attempt to quantify the reliability of ITDs, which forms the foundation for the rest of the study. However, it is not entirely clear whether the method used for calculating ITD reliability is the most appropriate, and the way the data are presented raises a number of questions.
Why is IPD variability plotted instead of ITD variability (or indeed spatial reliability)? The relationship between these measures is likely to vary across frequency, which makes it difficult to compare ITD variability across frequency when IPDs are plotted. Normalizing data across frequencies also makes it difficult to compare different locations and acoustical conditions. For example, in Fig.1a and Fig.1b, the data shown for 3 kHz at ~160 degrees seems quantitatively and visually quite different, but the difference (in Fig.1c) appears to be negligible.
How well do the measures of ITD reliability used reflect real-world listening? For example, the model used to calculate ITD reliability appears to assume the same (flat) spectral profile for targets and distractors, which are presented simultaneously with the same temporal envelope, and a uniform spatial distribution of sounds across space. It is therefore unclear how robust the study's results are to violations of these assumptions.
Does facial ruff removal produce an isolated effect on ITD variability or does it also produce changes in directional gain, and the relationship between spatial cues and sound location? Although the study considers this issue in some places (e.g. Fig.2, Fig.5), a clearer presentation of the acoustical effects of facial ruff removal and their implications (for all locations, not just those to the front), as well as an attempt to understand how these acoustical changes lead to the observed changes in ITD reliability, would greatly strengthen the study. In addition, Fig.1 shows average ITD reliability across owls, but it would be helpful to know how consistent these measures are across owls, given individual variability in Head-Related Transfer Functions (HRTFs). This potentially has implications for the electrophysiological experiments, if the HRTFs of those animals were not measured. One specific question that is potentially very relevant is whether the facial ruff attenuates sounds presented behind the animal and whether it does so in a frequency-dependent way. In addition, if facial ruff removal enables ILDs to be used for azimuth, then ITDs may also become less necessary at higher frequencies, even if their reliability remains unchanged.
ELECTROPHYSIOLOGY: The electrophysiological recordings in young owls are impressive, particularly since they were done longitudinally (although the follow-up data in adults is not shown). The decision to look at the relationship between different tuning properties following different types of developmental experience (e.g. relationship between best ITD and best frequency in the absence/presence of a fully developed facial ruff) is also a major strength, particularly in light of the very interesting results observed. The authors have succeeded in identifying clear evidence for the importance of acoustical input for determining frequency-tuning properties in the auditory midbrain. However, a number of points remain unclear.
It is unclear why some analyses (Fig.5, Fig.7) are focused on frontal locations and frontally-tuned neurons. It is also unclear why neurons with a best ITDs of 0 are described as frontally tuned since locations behind the animal produce an ITD of 0 also. Related to this, in Fig.1, facial ruff removal appears to reduce IPD variability at low frequencies for locations to the rear (~160 degrees), where the ITD is likely to be close to 0. Neurons with a best ITD of 0 might therefore be expected to adjust their frequency tuning in opposite directions depending on whether they are tuned to frontal or rearward locations.
The study suggests that information about high-frequency ITDs is not passed on to the ICX if the ICX does not contain neurons that have a high best frequency. However, neurons might be sensitive to ITDs at frequencies other than the best frequency, particularly if their frequency tuning is broader. It is also unclear whether the best frequency of a neuron always corresponds to the frequency that provides the most reliable ITD information, which the study implicitly assumes.
-
